You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- CH 06Document19 pagesCH 06cyberyeung0% (1)
- Gastrointestinal Physiology MOSBYDocument309 pagesGastrointestinal Physiology MOSBYUlquiorra SchifferNo ratings yet
- Carolina Rabbit Dissection Guide PDFDocument26 pagesCarolina Rabbit Dissection Guide PDFclairosusNo ratings yet
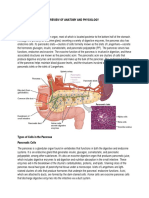
- Review of Anatomy and PhysiologyDocument7 pagesReview of Anatomy and PhysiologyKyla CalzadoNo ratings yet
- Clinical Manifestations and Diagnosis of Acute Pancreatitis PDFDocument32 pagesClinical Manifestations and Diagnosis of Acute Pancreatitis PDFChumas VidalNo ratings yet
- BECE Integrated Science Past QuestionsDocument20 pagesBECE Integrated Science Past QuestionsAnonymous lnC6IDg100% (4)
- JBDS Oncology Guideline Final Revised September 2021Document57 pagesJBDS Oncology Guideline Final Revised September 2021Majid KhanNo ratings yet
- Necrotizing Pancreatitis - DBarilDocument25 pagesNecrotizing Pancreatitis - DBarilataner1991No ratings yet
- Case Write Up Acute PancreatitisDocument25 pagesCase Write Up Acute PancreatitislunaNo ratings yet
- A Biased Look at BiomarkersDocument50 pagesA Biased Look at Biomarkerssiddu76No ratings yet
- Bifenil PolicloruratiDocument64 pagesBifenil PolicloruratiDoria IulianaNo ratings yet
- WaterDocument0 pagesWaterDoria IulianaNo ratings yet
- PesticidesDocument62 pagesPesticidesDoria IulianaNo ratings yet
- C VasileDocument15 pagesC VasileDoria IulianaNo ratings yet
- DM Reporting ZDocument52 pagesDM Reporting ZZsazsaNo ratings yet
- Digestive Organs & Their HistologyDocument5 pagesDigestive Organs & Their HistologyMall HiNo ratings yet
- Introduction to the Human Digestive SystemDocument45 pagesIntroduction to the Human Digestive SystemZakia Hasan ThalibNo ratings yet
- Diagnosis and Management of Insulinoma PDFDocument10 pagesDiagnosis and Management of Insulinoma PDFVanessa YunqueNo ratings yet
- 1st Term s3 BiologyDocument35 pages1st Term s3 BiologyAdio Babatunde Abiodun CabaxNo ratings yet
- Signs and Symptoms of Pancreatic Cancer Fact Sheet Dec 2014Document6 pagesSigns and Symptoms of Pancreatic Cancer Fact Sheet Dec 2014Nus EuNo ratings yet
- The Metabolic Solution Ebook PDFDocument26 pagesThe Metabolic Solution Ebook PDFCynthia WiikNo ratings yet
- Chapter 22 - Chemical Coordination and IntegrationDocument4 pagesChapter 22 - Chemical Coordination and IntegrationanuminiatureNo ratings yet
- Full Download Advanced Nutrition and Human Metabolism 5th Edition Gropper Test BankDocument35 pagesFull Download Advanced Nutrition and Human Metabolism 5th Edition Gropper Test Bankjeissyskylan100% (40)
- Biology Chapter 6 (6.4)Document34 pagesBiology Chapter 6 (6.4)Shirmei WangNo ratings yet
- Аcute and chronic pancreatitisDocument14 pagesАcute and chronic pancreatitisمیاں منیب احمدNo ratings yet
- JGCA JPN Classification 3rd EngDocument12 pagesJGCA JPN Classification 3rd EngLiz PesantesNo ratings yet
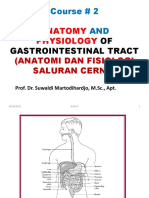
- Kuliah Ii - Blok 9 - Anatomi Dan Fisiologi Saluran CernaDocument54 pagesKuliah Ii - Blok 9 - Anatomi Dan Fisiologi Saluran Cernanovita istiqomahNo ratings yet
- Acute Pancreatitis 2023Document25 pagesAcute Pancreatitis 2023raphael chidiebereNo ratings yet
- CH 67Document14 pagesCH 67john mwangiNo ratings yet
- Drkamalkv 1 Ans June-14Document13 pagesDrkamalkv 1 Ans June-14Akshay MedkarNo ratings yet
- SGD Gross AnatomyDocument11 pagesSGD Gross Anatomyluis_chubeeNo ratings yet
- Review of Med Surg Exam QuestionsDocument30 pagesReview of Med Surg Exam QuestionsAntonella Vitale100% (3)
- Assignment 2 Austin InsleyDocument17 pagesAssignment 2 Austin Insleyapi-535501316No ratings yet
- Ch15 Metabolism of Dietary LipidsDocument37 pagesCh15 Metabolism of Dietary LipidsWahab KhaniNo ratings yet
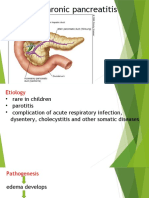
- Jurnal Pankreatitis KronisDocument7 pagesJurnal Pankreatitis KronisSalvia WatiNo ratings yet