You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- Logan SchemaDocument99 pagesLogan Schemamithos25678% (60)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 1 S NitrozilareDocument11 pages1 S NitrozilaredodutaNo ratings yet
- 3 Mecanisme Enzimatice NitrozilareDocument22 pages3 Mecanisme Enzimatice NitrozilaredodutaNo ratings yet
- A Digital Thermometer or Talk I2C To Your Atmel MicrocontrollerDocument7 pagesA Digital Thermometer or Talk I2C To Your Atmel MicrocontrollerdodutaNo ratings yet
- 3 Mecanisme Enzimatice NitrozilareDocument22 pages3 Mecanisme Enzimatice NitrozilaredodutaNo ratings yet
- NxtPhaseTD2003 RahmatianChavezDocument4 pagesNxtPhaseTD2003 RahmatianChavezdodutaNo ratings yet
- 12 dbSNODocument5 pages12 dbSNOdodutaNo ratings yet
- 10 Terapie S-NODocument10 pages10 Terapie S-NOdodutaNo ratings yet
- Arbotix Manual Draft2Document8 pagesArbotix Manual Draft2chauhanrishabh100% (1)
- Integrated Electronic Metering Insulator For Medium Voltage Overhead LinesDocument4 pagesIntegrated Electronic Metering Insulator For Medium Voltage Overhead LinesdodutaNo ratings yet
- Alstom Cosi NXCTDocument4 pagesAlstom Cosi NXCTdodutaNo ratings yet
- Integrated Electronic Metering Insulator For Medium Voltage Overhead LinesDocument4 pagesIntegrated Electronic Metering Insulator For Medium Voltage Overhead LinesdodutaNo ratings yet
- 2014 01 02 AdvancedOpenSSHDocument23 pages2014 01 02 AdvancedOpenSSHdodutaNo ratings yet
- Precision - Divider 4860Document6 pagesPrecision - Divider 4860dodutaNo ratings yet
- 3G-GSM Tactical Interception - Target LocationDocument51 pages3G-GSM Tactical Interception - Target Locationdoduta100% (2)
- 2014 01 02 AdvancedOpenSSHDocument23 pages2014 01 02 AdvancedOpenSSHdodutaNo ratings yet
- HOMEWORK 1 (General EnglishDocument4 pagesHOMEWORK 1 (General EnglishdodutaNo ratings yet
- Glossary of Legal TermsDocument46 pagesGlossary of Legal TermsdodutaNo ratings yet
- MC78XX/LM78XX/MC78XXA: 3-Terminal 1A Positive Voltage RegulatorDocument28 pagesMC78XX/LM78XX/MC78XXA: 3-Terminal 1A Positive Voltage Regulatorapi-19786583No ratings yet
- ICM7555, ICM7556: General Purpose Timers FeaturesDocument11 pagesICM7555, ICM7556: General Purpose Timers FeaturesdodutaNo ratings yet
- DNA Technology and The Human Genome WorksheetDocument4 pagesDNA Technology and The Human Genome WorksheetEamon BarkhordarianNo ratings yet
- Erythropoiesis Stages and FactorsDocument13 pagesErythropoiesis Stages and FactorsEng CirroNo ratings yet
- Southern Blotting: PrincipleDocument6 pagesSouthern Blotting: PrincipleArin AliNo ratings yet
- Laboratory Activity #1 The Cell (1) (1) 2Document3 pagesLaboratory Activity #1 The Cell (1) (1) 2Kim VenturaNo ratings yet
- Amino Acids Peptides and Proteins FinalDocument72 pagesAmino Acids Peptides and Proteins FinalHEMA 22No ratings yet
- Understanding Enzymes Samplepages-Revised PDFDocument62 pagesUnderstanding Enzymes Samplepages-Revised PDFAbe LimNo ratings yet
- Lahore University Biophysical Methods CourseDocument5 pagesLahore University Biophysical Methods CourseAnonymous sF8ZuiGNo ratings yet
- Biodegredation of UrethaneDocument8 pagesBiodegredation of UrethaneFuat TopuzNo ratings yet
- Repression and Activation of Promoter-Bound RNA Polymerase Activity by Gal RepressorDocument8 pagesRepression and Activation of Promoter-Bound RNA Polymerase Activity by Gal Repressoraxva1663No ratings yet
- Lab Report Cellular TransportDocument9 pagesLab Report Cellular Transportİlayda PınarNo ratings yet
- RNA Staining TechniquesDocument25 pagesRNA Staining TechniquesAvi VermaNo ratings yet
- Models of LifeDocument353 pagesModels of Lifeguillermo_feliúNo ratings yet
- Hexose Monophosphate ShuntDocument2 pagesHexose Monophosphate ShuntDr. SHIVA AITHAL100% (5)
- Ramesh Dnyandev KaleDocument2 pagesRamesh Dnyandev KaleSunil Chauhan-SathvaraNo ratings yet
- Sarthak Mechanism of Blood ClottingDocument42 pagesSarthak Mechanism of Blood ClottingVishal MishraNo ratings yet
- 7 E's Lesson 3 - ReplicationDocument7 pages7 E's Lesson 3 - Replicationcliff mascardo0% (1)
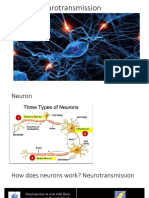
- Neurotransmission Processes and ReceptorsDocument66 pagesNeurotransmission Processes and ReceptorsSheik MohammedNo ratings yet
- Pepsin Enzyme Activity LabDocument4 pagesPepsin Enzyme Activity LabDebrah DebbieNo ratings yet
- VERI-Q™ PCR 316 Detection Platform: COVID-19Document2 pagesVERI-Q™ PCR 316 Detection Platform: COVID-19Smithesh TpNo ratings yet
- Cambridge International AS & A Level: Biology October/November 2022 2 HoursDocument28 pagesCambridge International AS & A Level: Biology October/November 2022 2 HourshudaNo ratings yet
- Chapter On TranscriptomicsDocument13 pagesChapter On TranscriptomicsMazhar FarNo ratings yet
- Sys Bio TextDocument25 pagesSys Bio TextAlpahNo ratings yet
- Eukaryotic DNA PolymerasesDocument8 pagesEukaryotic DNA PolymerasesvinodkumarmishraNo ratings yet
- Osteoimmunology in Orthodontic Tooth MovementDocument11 pagesOsteoimmunology in Orthodontic Tooth Movementdaniela2201No ratings yet
- FINAL KZN Grade 12 March 2020 Life SciencesDocument14 pagesFINAL KZN Grade 12 March 2020 Life SciencesAmahleNo ratings yet
- Phytohormones and Their FunctionsDocument85 pagesPhytohormones and Their FunctionsSuvrankar Datta100% (2)
- MCQ 1. in Transcription,: Type of Question: Content FromDocument2 pagesMCQ 1. in Transcription,: Type of Question: Content FromAlex XanderNo ratings yet
- DNA LibrariesDocument37 pagesDNA LibrariesMansoor KhalidNo ratings yet
- Isolation and Qualitative Analysis of Nucleic Acids (DNA From Onion)Document3 pagesIsolation and Qualitative Analysis of Nucleic Acids (DNA From Onion)Elina Lantion100% (1)
- CH 12 Reading GuideDocument9 pagesCH 12 Reading GuideKapil NathanNo ratings yet