You might also like
- Zong Yi Hu Et Al - Neurosteroids: Oligodendrocyte Mitochondria Convert Cholesterol To PregnenoloneDocument5 pagesZong Yi Hu Et Al - Neurosteroids: Oligodendrocyte Mitochondria Convert Cholesterol To PregnenoloneLonkesNo ratings yet
- Dahlqvist1968 - Alfa Glucosidase OkDocument9 pagesDahlqvist1968 - Alfa Glucosidase OkLeon NikolaiNo ratings yet
- Methods: A Comparison of Three of Glycogen Measurement in TissuesDocument8 pagesMethods: A Comparison of Three of Glycogen Measurement in TissuesDawn KentNo ratings yet
- Pengaruh Ekstrak Metanol Biji Mahoni (Swetenia Mahagoni Jacq) Terhadap Penurunan Glukosa Darah Dan Perbaikan Jaringan Pankreas Tikus Hasil Induksi MLD-STZDocument10 pagesPengaruh Ekstrak Metanol Biji Mahoni (Swetenia Mahagoni Jacq) Terhadap Penurunan Glukosa Darah Dan Perbaikan Jaringan Pankreas Tikus Hasil Induksi MLD-STZlailatul rofiahNo ratings yet
- Mi 06 201 PDFDocument10 pagesMi 06 201 PDFSavio PitombeiraNo ratings yet
- A New Method To Obtain B-Glucan From Saccharomyces Cerevisiae CellsDocument4 pagesA New Method To Obtain B-Glucan From Saccharomyces Cerevisiae Cellshuynhtanvu2011No ratings yet
- Perifusion Steroidogenesis Leydig: LH-dependentDocument9 pagesPerifusion Steroidogenesis Leydig: LH-dependentJean Pierre Chastre LuzaNo ratings yet
- GliclazideDocument8 pagesGliclazideUpe VeerlapatiNo ratings yet
- Tuna 5Document9 pagesTuna 5Leonardo SánchezNo ratings yet
- Research SampleDocument11 pagesResearch SamplesuryasanNo ratings yet
- Sterol Carrier Protein and Fatty Acid-Binding ProteinDocument7 pagesSterol Carrier Protein and Fatty Acid-Binding ProteinMartin OreNo ratings yet
- Research Article: Increased Yield and Improved Transplantation Outcome of Mouse Islets With Bovine Serum AlbuminDocument10 pagesResearch Article: Increased Yield and Improved Transplantation Outcome of Mouse Islets With Bovine Serum AlbuminCLPHtheoryNo ratings yet
- Biochemical Properties of A B-Xylosidase From: Clostridium CellulolyticumDocument4 pagesBiochemical Properties of A B-Xylosidase From: Clostridium CellulolyticumPedro HamannNo ratings yet
- 1988 - Microencapsulation of Pancreatic Islet Cells - A Bioartificial Endocrine PancreasDocument6 pages1988 - Microencapsulation of Pancreatic Islet Cells - A Bioartificial Endocrine PancreasAndrés MenesesNo ratings yet
- Invertase Lab 1 and 2Document8 pagesInvertase Lab 1 and 2Tiyah TimothyNo ratings yet
- Invertase Lab 1 or 2Document8 pagesInvertase Lab 1 or 2Tiyah TimothyNo ratings yet
- Research Paper CholesterolDocument18 pagesResearch Paper CholesterolTony NgNo ratings yet
- Cellulase Enzyme Production by Streptomyces SP Using Fruit Waste As SubstrateDocument5 pagesCellulase Enzyme Production by Streptomyces SP Using Fruit Waste As SubstrateHamka NurkayaNo ratings yet
- DPPH Radical-Scavenging Compounds From Dou-Chi, A Soybean Fermented FoodDocument0 pagesDPPH Radical-Scavenging Compounds From Dou-Chi, A Soybean Fermented FoodEllen SantosNo ratings yet
- Supporting Online Material For: Regulation of Cellular Metabolism by Protein Lysine AcetylationDocument39 pagesSupporting Online Material For: Regulation of Cellular Metabolism by Protein Lysine AcetylationEdmond MengoueNo ratings yet
- Obestatina y Los Cambios en Los AdipositosDocument9 pagesObestatina y Los Cambios en Los AdipositosDavid AriasNo ratings yet
- In Final Form May 6, 1999)Document10 pagesIn Final Form May 6, 1999)CLPHtheoryNo ratings yet
- Mutanolisina EnglishDocument10 pagesMutanolisina EnglishCristobal Hernandez ToledoNo ratings yet
- Wang2012 Article TheEffectOfIntracellularAminoADocument8 pagesWang2012 Article TheEffectOfIntracellularAminoALadysNo ratings yet
- Email: Humphreysa001@mymail - Mssu.edu Genetics (BIO305) Spring 2023Document14 pagesEmail: Humphreysa001@mymail - Mssu.edu Genetics (BIO305) Spring 2023api-721737889No ratings yet
- Glycolysis-Deficient: Studies Cells That Production of TumorDocument5 pagesGlycolysis-Deficient: Studies Cells That Production of TumorFabian BalderrabanoNo ratings yet
- 10.1016@0006 2952 (85) 90529 5Document6 pages10.1016@0006 2952 (85) 90529 5BoengkerNo ratings yet
- Mutation of An Inosine Producing Strain of Bacillus Subtilis For Guanosine ProductionDocument5 pagesMutation of An Inosine Producing Strain of Bacillus Subtilis For Guanosine Production김영우No ratings yet
- Dandang Gendis (Clinacanthus Nutans PDFDocument6 pagesDandang Gendis (Clinacanthus Nutans PDFnatinlalaNo ratings yet
- How Noscapine Metabolise Heme?Document15 pagesHow Noscapine Metabolise Heme?Chief editorNo ratings yet
- Coffee Pulp Waste Substrate Based in Cellulase Production by Penicillium Sp. VT11 Under Solid-State FermentationDocument9 pagesCoffee Pulp Waste Substrate Based in Cellulase Production by Penicillium Sp. VT11 Under Solid-State FermentationNabilah Ilmalah SunartoNo ratings yet
- Adenovirus Methods and ProtocolsDocument340 pagesAdenovirus Methods and ProtocolsdurerosNo ratings yet
- In Vitro & in Vivo Estrogenic Activity of Glycoside Fractions of Solanum Nigrum FruitDocument6 pagesIn Vitro & in Vivo Estrogenic Activity of Glycoside Fractions of Solanum Nigrum FruitKocyd Dyck OkooNo ratings yet
- Fungal EnzymesDocument9 pagesFungal EnzymesYui Bioscience PalsanNo ratings yet
- Ginger PublishedDocument8 pagesGinger PublishedFatimoh AbdulsalamNo ratings yet
- 1 PBDocument6 pages1 PBUncu EchiNo ratings yet
- LactosacarosaDocument8 pagesLactosacarosaAriel plutarco cardoso herreraNo ratings yet
- Momordica Charantia Fruit Juice Stimulates: Glucose and Amino Acid Uptakes in L6 MyotubesDocument6 pagesMomordica Charantia Fruit Juice Stimulates: Glucose and Amino Acid Uptakes in L6 MyotubesEliani BazanNo ratings yet
- Insulin Secretion From Perifused Rat Pancreatic PseudoisletsDocument7 pagesInsulin Secretion From Perifused Rat Pancreatic PseudoisletskalwusaidiNo ratings yet
- Nicotine Induced Oxidative Damage in Rat Lymphocytes - An: InvitrostudyDocument4 pagesNicotine Induced Oxidative Damage in Rat Lymphocytes - An: InvitrostudyIOSR Journal of PharmacyNo ratings yet
- A New Protocol For Isolating Cardiac Myocytes Used in Patch-Clamp StudiesDocument12 pagesA New Protocol For Isolating Cardiac Myocytes Used in Patch-Clamp StudiesSathya DevNo ratings yet
- Amylolytic Enzymes Produced by B. Amyloliquefaciens MIR41Document6 pagesAmylolytic Enzymes Produced by B. Amyloliquefaciens MIR41Guillermo Raul CastroNo ratings yet
- Research Article Boletus SpeciosusDocument8 pagesResearch Article Boletus SpeciosusCamilo BautistaNo ratings yet
- 6 Sy Comparative Evaluation of Niosome Formulations Prepared by Different TechniquesDocument6 pages6 Sy Comparative Evaluation of Niosome Formulations Prepared by Different Techniquesgrvgr8No ratings yet
- Foods: Gluten Detection and Speciation by Liquid Chromatography Mass Spectrometry (LC-MS/MS)Document17 pagesFoods: Gluten Detection and Speciation by Liquid Chromatography Mass Spectrometry (LC-MS/MS)giamilliaNo ratings yet
- 3331 FullDocument7 pages3331 FullSukhvinder Singh RanaNo ratings yet
- Antidiabetic Activity of Ruellia Tuberosa L Role oDocument9 pagesAntidiabetic Activity of Ruellia Tuberosa L Role orandy hidayatNo ratings yet
- FulltextDocument5 pagesFulltextAmeba OioNo ratings yet
- Studies of Thrombolytic, Antioxidant and Cytotoxic Properties of Prunus DomesticaDocument3 pagesStudies of Thrombolytic, Antioxidant and Cytotoxic Properties of Prunus DomesticaMohammad Shohel RanaNo ratings yet
- Neuorospora CrassaDocument4 pagesNeuorospora CrassaAleya NasreenNo ratings yet
- Biocompatibility of Graphene OxideDocument8 pagesBiocompatibility of Graphene OxidelpuresearchNo ratings yet
- Plan Cience: Isolation and Properties of A Lectin From The Seeds ofDocument6 pagesPlan Cience: Isolation and Properties of A Lectin From The Seeds ofRana KameiNo ratings yet
- Validation of The Analytical Method For Determination of Meloxicam and Bioequivalence Study From Meloxicam Containing Microparticle Formulations in RabbitsDocument16 pagesValidation of The Analytical Method For Determination of Meloxicam and Bioequivalence Study From Meloxicam Containing Microparticle Formulations in RabbitsBerlian Rizky PutraNo ratings yet
- Micro Sphere ThesisDocument26 pagesMicro Sphere ThesisPeter SamNo ratings yet
- Cytotoxicity: in Vitro Determination: Cell CulturesDocument2 pagesCytotoxicity: in Vitro Determination: Cell CulturesUday BaruahNo ratings yet
- Altas Presiones - Promover Peptidos Bioactivos OvoalbúminaDocument6 pagesAltas Presiones - Promover Peptidos Bioactivos Ovoalbúminamdsanchezo8373No ratings yet
- Bacterii Celulozolitice PDFDocument6 pagesBacterii Celulozolitice PDFArisitica VinilescuNo ratings yet
- Live Cell Assays: From Research to Regulatory ApplicationsFrom EverandLive Cell Assays: From Research to Regulatory ApplicationsNo ratings yet
- Oligonucleotide-Based Drugs and Therapeutics: Preclinical and Clinical Considerations for DevelopmentFrom EverandOligonucleotide-Based Drugs and Therapeutics: Preclinical and Clinical Considerations for DevelopmentNicolay FerrariNo ratings yet
- WHO Diabetes Definition and DiagnosisDocument50 pagesWHO Diabetes Definition and DiagnosisDavidNo ratings yet
- QB 060100302Document2 pagesQB 060100302Haleema SultanNo ratings yet
- Animal Models For Anti Diabetic ActivityDocument11 pagesAnimal Models For Anti Diabetic ActivityHaleema SultanNo ratings yet
- Polymerase Chain Reaction (PCR)Document37 pagesPolymerase Chain Reaction (PCR)Haleema SultanNo ratings yet
- Screening of Anticonvulsant DrugsDocument14 pagesScreening of Anticonvulsant DrugsHaleema SultanNo ratings yet
- Animal Rights: "The Worst Sin Towards Our Fellow Creatures Is Not To Hate Them, But ToDocument22 pagesAnimal Rights: "The Worst Sin Towards Our Fellow Creatures Is Not To Hate Them, But ToHaleema SultanNo ratings yet
- Interpreting Infrared Spectra Painlessly, Quickly, and CorrectlyDocument5 pagesInterpreting Infrared Spectra Painlessly, Quickly, and CorrectlyHaleema SultanNo ratings yet
- Myocardial Ischemia: Dr. Wael H. Mansy, MDDocument37 pagesMyocardial Ischemia: Dr. Wael H. Mansy, MDHaleema SultanNo ratings yet
- Screening of Anti-Inflammatory DrugsDocument11 pagesScreening of Anti-Inflammatory DrugsHaleema SultanNo ratings yet
- Institute of Pharmacy Nirma University: Admission Processing Fee (Including Form Fee) : Rs. 1,000/-Web Downloaded FormDocument5 pagesInstitute of Pharmacy Nirma University: Admission Processing Fee (Including Form Fee) : Rs. 1,000/-Web Downloaded FormHaleema SultanNo ratings yet
- Interpretation of Infrared Spectra, A Practical ApproachDocument24 pagesInterpretation of Infrared Spectra, A Practical ApproachLucas TimmerNo ratings yet
- Generic NameDocument34 pagesGeneric NameHaleema SultanNo ratings yet
- My Project ETHOSOMESDocument26 pagesMy Project ETHOSOMESHaleema SultanNo ratings yet
- Brain TumoursDocument35 pagesBrain TumoursHealth Education Library for PeopleNo ratings yet
- Zincum ValerianicumDocument3 pagesZincum ValerianicumKamalNo ratings yet
- Standard Operating Proceduresgov1Document197 pagesStandard Operating Proceduresgov1Adam ShawNo ratings yet
- Sphygmomanometer FileDocument12 pagesSphygmomanometer FileKye GarciaNo ratings yet
- Topic Outline: APR 14, 2021 Dr. Michelle BuelaDocument5 pagesTopic Outline: APR 14, 2021 Dr. Michelle BuelaSGD5Christine MendozaNo ratings yet
- G11 Biology EnglishDocument7 pagesG11 Biology Englishahmed aluNo ratings yet
- MHD Exam 6 MaterialDocument179 pagesMHD Exam 6 Materialnaexuis5467100% (1)
- ECG (Cardiology Clinic 2006)Document127 pagesECG (Cardiology Clinic 2006)VuqarNo ratings yet
- f3 Chapter 1Document110 pagesf3 Chapter 1miracleambitiousNo ratings yet
- The+O2+Awakening+ +O2+Daily+Practice+GuideDocument3 pagesThe+O2+Awakening+ +O2+Daily+Practice+GuidetonalbadassNo ratings yet
- Brain Slides SEMINAR 1 - 220606 - 142811 - 220606 - 223805Document32 pagesBrain Slides SEMINAR 1 - 220606 - 142811 - 220606 - 223805pang pangNo ratings yet
- Ebook Human Anatomy Color Atlas and Textbook PDF Full Chapter PDFDocument67 pagesEbook Human Anatomy Color Atlas and Textbook PDF Full Chapter PDFronald.mcneil866100% (23)
- Foreign Pharmacy Graduate Equivalency ExaminationDocument26 pagesForeign Pharmacy Graduate Equivalency Examinationnewfakeemail100% (2)
- Geeky Medics DocumentationDocument3 pagesGeeky Medics DocumentationGus LionsNo ratings yet
- Ecg en LabviewDocument61 pagesEcg en LabviewHarold David Gil MuñozNo ratings yet
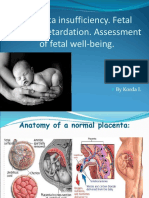
- Placenta InsufficiencyDocument39 pagesPlacenta InsufficiencyslyfoxkittyNo ratings yet
- Medtronic Physio Control Lifepak 10 DefibrillatorDocument2 pagesMedtronic Physio Control Lifepak 10 DefibrillatorCarlos RomeroNo ratings yet
- Cansuje National High School: Quarter 3 Summative Test No. 1 in Science 10Document3 pagesCansuje National High School: Quarter 3 Summative Test No. 1 in Science 10Jannet FuentesNo ratings yet
- (E BOOK) Martini Essentials of Anatomy PhysiologyDocument3 pages(E BOOK) Martini Essentials of Anatomy PhysiologyRaisa sya12No ratings yet
- Introduction To Molecular Biology and GeneticsDocument146 pagesIntroduction To Molecular Biology and GeneticsRedaGaafar100% (1)
- Food Tambola: Department of Food Science & TechnologyDocument183 pagesFood Tambola: Department of Food Science & TechnologyManu BhatiaNo ratings yet
- FFHP 6Document29 pagesFFHP 6Danny GoldstoneNo ratings yet
- FeverDocument12 pagesFeverJ.B. BuiNo ratings yet
- PathologyDocument36 pagesPathologySayeed KhanNo ratings yet
- Polymers in Medicine and Surgery 1975 PDFDocument336 pagesPolymers in Medicine and Surgery 1975 PDFVidvendu GuptaNo ratings yet
- Vulnus Wound HealingDocument20 pagesVulnus Wound HealingVicKy AmaliaNo ratings yet
- Craniofacial Biology Notes Unit 1Document37 pagesCraniofacial Biology Notes Unit 1nohacksNo ratings yet
- Notes - Human GeneticsDocument68 pagesNotes - Human GeneticsiwennieNo ratings yet
- Animal Tissue Culture: Ms. Veena ShriramDocument38 pagesAnimal Tissue Culture: Ms. Veena Shriramw5waNo ratings yet
- An Estimation of The Number of Cells in The Human BodyDocument11 pagesAn Estimation of The Number of Cells in The Human BodysedilotNo ratings yet