You might also like
- 9D NLBA Illustrated Glossary 2015Document141 pages9D NLBA Illustrated Glossary 2015galante81No ratings yet
- Blood Circulation System 10 PDFDocument29 pagesBlood Circulation System 10 PDFSardar jiNo ratings yet
- Overview of Cardiovascular System: An Introduction To Chapters 9 - 24 and Chapter 36 Guyton and Hall, 12 EditionDocument10 pagesOverview of Cardiovascular System: An Introduction To Chapters 9 - 24 and Chapter 36 Guyton and Hall, 12 EditionbahahahahNo ratings yet
- Electrocardiography - 2Document17 pagesElectrocardiography - 2salochinNo ratings yet
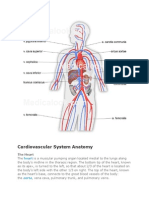
- Cardiovascular System AnatomyDocument16 pagesCardiovascular System AnatomyJona Addatu0% (1)
- A Model of Human Postural Control Inspired by Separated Human Sensory SystemsDocument10 pagesA Model of Human Postural Control Inspired by Separated Human Sensory SystemspamelaNo ratings yet
- Central Venous Pressure: Its Clinical Use and Role in Cardiovascular DynamicsFrom EverandCentral Venous Pressure: Its Clinical Use and Role in Cardiovascular DynamicsNo ratings yet
- Guidelines For Registration As A Professional Engineer - Peb SingaporeDocument9 pagesGuidelines For Registration As A Professional Engineer - Peb SingaporejeffreymacaseroNo ratings yet
- 3 - Tips For Professional Engineer Interview in MalaysiaDocument19 pages3 - Tips For Professional Engineer Interview in Malaysiazahrul azha100% (8)
- Pump Handbook by GrundfossDocument156 pagesPump Handbook by GrundfossSyam Tawakkal100% (1)
- CP On Pre-EclampsiaDocument152 pagesCP On Pre-Eclampsiakathy100% (2)
- Steel Beam BS 5950Document17 pagesSteel Beam BS 5950Vashish RamrechaNo ratings yet
- Max Allowable Pressure of Pipes and Pipellines CER 04022013 1 (V. S. Kumar Unprotected)Document106 pagesMax Allowable Pressure of Pipes and Pipellines CER 04022013 1 (V. S. Kumar Unprotected)Vashish Ramrecha100% (1)
- Complexity and HumanDocument25 pagesComplexity and Humanraku2121No ratings yet
- IISRT Jaisre Kamaraj (EC)Document5 pagesIISRT Jaisre Kamaraj (EC)IISRTNo ratings yet
- Determinants of Cardiac Function SimulationDocument23 pagesDeterminants of Cardiac Function SimulationDavid MercadoNo ratings yet
- Eletromecanichal HeartDocument5 pagesEletromecanichal HeartAntonioSouzaNo ratings yet
- Markers of Altered Cardiovascular Autonomic Function in Overweight ChildrenDocument4 pagesMarkers of Altered Cardiovascular Autonomic Function in Overweight Childrenshanto shahrinNo ratings yet
- Simulation of The Cardiovascular Mechanical System Based On Pressure-Flow Model Rest ConditionDocument12 pagesSimulation of The Cardiovascular Mechanical System Based On Pressure-Flow Model Rest ConditionInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Bondgraph Published CardioVascular 1Document4 pagesBondgraph Published CardioVascular 1AhmedNo ratings yet
- Physiological Modeling - Final Project (Circulatory System)Document18 pagesPhysiological Modeling - Final Project (Circulatory System)Joshua DamianNo ratings yet
- Human Heart Modelling Using Hydro Electro Mechanical SystemsDocument25 pagesHuman Heart Modelling Using Hydro Electro Mechanical Systemsmaggi3012No ratings yet
- Reserch VestDocument11 pagesReserch VestRAMO STEF SZEKERESNo ratings yet
- Modelling of The Respiratory System For PDFDocument8 pagesModelling of The Respiratory System For PDFshipraNo ratings yet
- Cardiovascular Model With Human Elastance Function and Valve DynamicsDocument7 pagesCardiovascular Model With Human Elastance Function and Valve DynamicsAndreea Diana GhitaNo ratings yet
- Dynamical Compensation in Physiological Circuits: Omer Karin, Avital Swisa, Benjamin Glaser, Yuval Dor & Uri AlonDocument7 pagesDynamical Compensation in Physiological Circuits: Omer Karin, Avital Swisa, Benjamin Glaser, Yuval Dor & Uri AlonFrancisco Vicent PachecoNo ratings yet
- Guyton The Surprising Kidney Fluid Mechanism For Pressure Control Its Infinite GainDocument7 pagesGuyton The Surprising Kidney Fluid Mechanism For Pressure Control Its Infinite GainAryNo ratings yet
- ProposalDocument2 pagesProposaltoddnkNo ratings yet
- Time Delays, Oscillations, and Chaos in Physiological Control SystemsDocument15 pagesTime Delays, Oscillations, and Chaos in Physiological Control SystemsFrancis TomyNo ratings yet
- Physiological Modeling, Simulation, and Control: Joseph L. PalladinoDocument30 pagesPhysiological Modeling, Simulation, and Control: Joseph L. PalladinoqhqhqNo ratings yet
- Pumping MechanismsDocument10 pagesPumping MechanismsfuggaNo ratings yet
- Biocybernetics: By: Fragoon M. Ahmed, PHDDocument29 pagesBiocybernetics: By: Fragoon M. Ahmed, PHDBkry BosatyNo ratings yet
- 03C Modeling A Heart Pump PDFDocument20 pages03C Modeling A Heart Pump PDFMarlon Andres Navarro AlvarezNo ratings yet
- Mathematical and Physical Models of A Total Artificial HeartDocument6 pagesMathematical and Physical Models of A Total Artificial HeartBishal UchilNo ratings yet
- Zhoujian Cao Et Al - Turbulence Control With Local Pacing and Its Implication in Cardiac DefibrillationDocument9 pagesZhoujian Cao Et Al - Turbulence Control With Local Pacing and Its Implication in Cardiac DefibrillationGretymjNo ratings yet
- Biocybernetics: Dr. Fragoon AhmedDocument28 pagesBiocybernetics: Dr. Fragoon AhmedBkry BosatyNo ratings yet
- Computers in Biology and Medicine: J. Fernandez de Canete, J. Luque, J. Barbancho, V. MunozDocument9 pagesComputers in Biology and Medicine: J. Fernandez de Canete, J. Luque, J. Barbancho, V. MunozGerai ZahraNo ratings yet
- Permatasari 2018 J. Phys. Conf. Ser. 1025 012086Document8 pagesPermatasari 2018 J. Phys. Conf. Ser. 1025 012086deepNo ratings yet
- Tolerancia y Dependencia de DrogasDocument10 pagesTolerancia y Dependencia de DrogasLopez LionelNo ratings yet
- Computational Principles of Movement NeuroscienceDocument6 pagesComputational Principles of Movement NeuroscienceNicolas Espinoza ConstanzoNo ratings yet
- Bondgraph Published CardioVascular 2.1Document18 pagesBondgraph Published CardioVascular 2.1AhmedNo ratings yet
- Open Loop and Close Loop System 1Document11 pagesOpen Loop and Close Loop System 1Bhuban LimbuNo ratings yet
- Cooper 2005Document12 pagesCooper 20050hitk0No ratings yet
- Circulatory Physiology III: Arterial Pressure and HypertensionDocument20 pagesCirculatory Physiology III: Arterial Pressure and Hypertensionraanja2No ratings yet
- 19 Vol 92 No 1Document9 pages19 Vol 92 No 1lahlouhNo ratings yet
- Accepted Manuscript: 10.1016/j.physbeh.2018.06.008Document43 pagesAccepted Manuscript: 10.1016/j.physbeh.2018.06.008Carolina MarroquinNo ratings yet
- Physiology-1st Year Topical Past Papers-1Document14 pagesPhysiology-1st Year Topical Past Papers-1HussnainNo ratings yet
- Control of Leg Movements Driven by Electrically Stimulated MusclesDocument7 pagesControl of Leg Movements Driven by Electrically Stimulated MusclesBounty JhaNo ratings yet
- Comparison of Nonlinear Mammalian Cochlear-Partition ModelsDocument42 pagesComparison of Nonlinear Mammalian Cochlear-Partition ModelsRobert SzalaiNo ratings yet
- Human Anatomy and Physio Revision NotesDocument3 pagesHuman Anatomy and Physio Revision Noteshuangc17No ratings yet
- Hypertension: A Comparative Review Based On Fractal Wave Theory of ContinuumDocument8 pagesHypertension: A Comparative Review Based On Fractal Wave Theory of ContinuumAdaptive MedicineNo ratings yet
- Bondgraph Published CardioVascular 5Document24 pagesBondgraph Published CardioVascular 5AhmedNo ratings yet
- Timothy J. Lewis and Michael R. Guevara - Chaotic Dynamics in An Ionic Model of The Propagated Cardiac Action PotentialDocument26 pagesTimothy J. Lewis and Michael R. Guevara - Chaotic Dynamics in An Ionic Model of The Propagated Cardiac Action PotentialLuammmNo ratings yet
- Paper of Class 4Document12 pagesPaper of Class 4Shreetama BhattacharyaNo ratings yet
- 1994 UrsinoDocument16 pages1994 UrsinosebasgoryNo ratings yet
- Gromacs Molecular Modeling TutorialDocument11 pagesGromacs Molecular Modeling Tutorialca_teixeiraNo ratings yet
- Melo2018 CV Adaptation by RTDocument8 pagesMelo2018 CV Adaptation by RTMumtaz MaulanaNo ratings yet
- 1996 Nonlinear Control of Heart Rate Variability in Human InfantsDocument6 pages1996 Nonlinear Control of Heart Rate Variability in Human InfantsSP507No ratings yet
- Revisiting The Simplified Bernoulli EquationDocument6 pagesRevisiting The Simplified Bernoulli Equationgu1000No ratings yet
- Physiology - 1st Year - Topical PapersDocument13 pagesPhysiology - 1st Year - Topical PapersKashmala WasiqNo ratings yet
- Muscle Mechanics and Reflexes Are Not Tuned For Disturbance RejectionDocument5 pagesMuscle Mechanics and Reflexes Are Not Tuned For Disturbance RejectionverdosNo ratings yet
- Introduction Tobio MechatronicsDocument31 pagesIntroduction Tobio MechatronicsEmreEmanetNo ratings yet
- Guytons Diagram Brought To Life From Graphic ChartDocument15 pagesGuytons Diagram Brought To Life From Graphic ChartAdib npmNo ratings yet
- XyzDocument100 pagesXyzAnonymous kpQrRs2NPNo ratings yet
- Human Gait Analysed by An Artificial Neural Network ModelDocument4 pagesHuman Gait Analysed by An Artificial Neural Network Modelstroci4047No ratings yet
- A Mathematical Model of Human Heart Including The Effects of Heart (2018)Document9 pagesA Mathematical Model of Human Heart Including The Effects of Heart (2018)Maria Paula Lopez UribeNo ratings yet
- Control Theory in Biology and Medicine: Introduction To The Special IssueDocument6 pagesControl Theory in Biology and Medicine: Introduction To The Special IssueWellington Cássio PinheiroNo ratings yet
- Condicionamiento Reflejos PosturalesDocument14 pagesCondicionamiento Reflejos PosturalesFrancisco Vicent PachecoNo ratings yet
- Equipment ListDocument10 pagesEquipment ListVashish RamrechaNo ratings yet
- 2017 1 Art 06 PDFDocument12 pages2017 1 Art 06 PDFG V LakshmiNo ratings yet
- Piping FormulaeDocument9 pagesPiping FormulaeVashish RamrechaNo ratings yet
- Steel Cals SimpleDocument53 pagesSteel Cals SimpleVashish RamrechaNo ratings yet
- DC CD Enteric Line List TemplateDocument1 pageDC CD Enteric Line List TemplateVashish RamrechaNo ratings yet
- Beam CantileverDocument16 pagesBeam CantileverAdam SlametNo ratings yet
- Gantt ChartDocument6 pagesGantt ChartVashish RamrechaNo ratings yet
- ISA SymbolsDocument1 pageISA SymbolsmustafasandikciNo ratings yet
- DC CD Enteric Line List TemplateDocument1 pageDC CD Enteric Line List TemplateVashish RamrechaNo ratings yet
- SLS in Manufacturing Sector-7thProofDocument90 pagesSLS in Manufacturing Sector-7thProofVashish RamrechaNo ratings yet
- Gantt ChartDocument6 pagesGantt ChartVashish RamrechaNo ratings yet
- In-Text: (Nickerson and Buist, 2008) Bibliography: Nickerson, D. and Buist, M. (2008) - Practical Application of Cellml 1.1: The IntegrationDocument1 pageIn-Text: (Nickerson and Buist, 2008) Bibliography: Nickerson, D. and Buist, M. (2008) - Practical Application of Cellml 1.1: The IntegrationVashish RamrechaNo ratings yet
- Steel Cals SimpleDocument53 pagesSteel Cals SimpleVashish RamrechaNo ratings yet
- 3.0 Strike Receivers AssessmentDocument5 pages3.0 Strike Receivers AssessmentVashish RamrechaNo ratings yet
- Roll 2015Document26 pagesRoll 2015Vashish RamrechaNo ratings yet
- Technical Report SynopsisDocument3 pagesTechnical Report SynopsisAzharuddin_kfupmNo ratings yet
- Edited FixturesDocument67 pagesEdited FixturesVashish RamrechaNo ratings yet
- Why Isometric Drawings Are RequiredDocument30 pagesWhy Isometric Drawings Are RequiredVashish RamrechaNo ratings yet
- Training Record Book 2011 PDFDocument25 pagesTraining Record Book 2011 PDFsarwarrosunNo ratings yet
- DSG MechDocument8 pagesDSG MechVashish RamrechaNo ratings yet
- CH-4 Shells Under External PressureDocument64 pagesCH-4 Shells Under External PressureVashish RamrechaNo ratings yet
- Legacy ECSA ProfEng Appform TESDocument1 pageLegacy ECSA ProfEng Appform TESVashish RamrechaNo ratings yet
- Legacy ECSA ProfEng Appform TERDocument1 pageLegacy ECSA ProfEng Appform TERVashish RamrechaNo ratings yet
- The Gurney Flap: A Case Study: 1.1 Issues in Wing DesignDocument9 pagesThe Gurney Flap: A Case Study: 1.1 Issues in Wing DesignVashish RamrechaNo ratings yet
- Fluid Management in Acute Kidney InjuryDocument8 pagesFluid Management in Acute Kidney InjuryHGZ 83 MoreliaNo ratings yet
- Pulmonary CirculationDocument36 pagesPulmonary CirculationSllavko K. KallfaNo ratings yet
- Quarter: 4 / Semester: 2 / WEEK: 4: Online Resource/sDocument17 pagesQuarter: 4 / Semester: 2 / WEEK: 4: Online Resource/sTristan Paul PagalananNo ratings yet
- Topic 9 Cardiovascular SystemDocument36 pagesTopic 9 Cardiovascular SystemAlejandra CastilloNo ratings yet
- Chapter 13 - Blood Vessels and CirculationDocument21 pagesChapter 13 - Blood Vessels and CirculationGabriel CastenNo ratings yet
- AQA Biol A Unit 1 Jan02Document20 pagesAQA Biol A Unit 1 Jan02Raymond RayNo ratings yet
- Quantitative Human Physiology An Introduction 1St Edition Feher Solutions Manual Full Chapter PDFDocument39 pagesQuantitative Human Physiology An Introduction 1St Edition Feher Solutions Manual Full Chapter PDFvernier.decyliclnn4100% (7)
- Respiration:: Mcqs PhysiologyDocument96 pagesRespiration:: Mcqs PhysiologyAhmedNo ratings yet
- Angiology: General Anatomy Dr. Sabah Rehman (MBBS, Mphil)Document51 pagesAngiology: General Anatomy Dr. Sabah Rehman (MBBS, Mphil)Ahsan IslamNo ratings yet
- MCQ TBL Cardiovascular Adaptation To Muscular ExerciseDocument40 pagesMCQ TBL Cardiovascular Adaptation To Muscular ExerciseAdlina Tajuddin100% (1)
- (Advances in Space Biology and Medicine 4) Sjoerd L. Bonting (Eds.) - Elsevier Science (1994)Document286 pages(Advances in Space Biology and Medicine 4) Sjoerd L. Bonting (Eds.) - Elsevier Science (1994)Sohnit KumarNo ratings yet
- Cardiovascular Control, Part 2: Bio Sci E109 Human PhysiologyDocument18 pagesCardiovascular Control, Part 2: Bio Sci E109 Human PhysiologyStephanie PinedaNo ratings yet
- A&P Exam 2 QuestionsDocument22 pagesA&P Exam 2 QuestionsAbdul QuorishyNo ratings yet
- Viruses: Endothelium Infection and Dysregulation by Sars-Cov-2: Evidence and Caveats in Covid-19Document26 pagesViruses: Endothelium Infection and Dysregulation by Sars-Cov-2: Evidence and Caveats in Covid-19Katie WinsNo ratings yet
- Flowing of Blood in HeartDocument5 pagesFlowing of Blood in HeartAkshay BahetyNo ratings yet
- Hypertension: Nucleus Medical MediaDocument17 pagesHypertension: Nucleus Medical Mediakrish oswalNo ratings yet
- 0000180Document53 pages0000180Nirmala MaranNo ratings yet
- Inotropic and Vasoactive Drugs in Pediatric ICUDocument6 pagesInotropic and Vasoactive Drugs in Pediatric ICUCandy RevolloNo ratings yet
- Cardiovascular SystemDocument17 pagesCardiovascular SystemPoojaNo ratings yet
- 7-Moto Renal - MCQsDocument62 pages7-Moto Renal - MCQspvs5155No ratings yet
- Chapter 21 - Blood Vessels and CirculationDocument13 pagesChapter 21 - Blood Vessels and CirculationSameer AkhtarNo ratings yet
- L2 PulmonaryCirculation PHAR6200 MMPH6172 173110Document85 pagesL2 PulmonaryCirculation PHAR6200 MMPH6172 173110Rankie ChoiNo ratings yet
- Pharm NotesDocument113 pagesPharm NotesBibek SutradharNo ratings yet
- VASODILATORSDocument2 pagesVASODILATORSMichiko_Lois_Tao_203No ratings yet
- FSG332 Cardiovascular Pathophysiology 3 - 2Document39 pagesFSG332 Cardiovascular Pathophysiology 3 - 2Huzaifa KhanNo ratings yet
- Kul Sistem CV Ut Prodi FarmasiDocument34 pagesKul Sistem CV Ut Prodi FarmasiMicho Zhii IntelNo ratings yet