You might also like
- Transporte CelularDocument8 pagesTransporte CelularFelipe Farfan100% (1)
- Guía Transporte Celular, 58Document7 pagesGuía Transporte Celular, 58Lisi GómezNo ratings yet
- Teoria CelularDocument65 pagesTeoria Celulartigres1212No ratings yet
- Celulas Eucariotas y ProcariotasDocument24 pagesCelulas Eucariotas y ProcariotasAle GallegosNo ratings yet
- Célula Eucariota y Célula ProcariotaDocument2 pagesCélula Eucariota y Célula ProcariotaMichael JonesNo ratings yet
- EnzimasDocument38 pagesEnzimaslisseth carboNo ratings yet
- Transporte y Relaciones - Membrana PDFDocument12 pagesTransporte y Relaciones - Membrana PDFBritney Charline AscherNo ratings yet
- Anatomia de La CelulaDocument22 pagesAnatomia de La CelulaBrady Rubén Sandoval Coveñas100% (3)
- Agua BioquímicaDocument99 pagesAgua BioquímicalupitagroNo ratings yet
- Metabolismo de Los CarbohidratosDocument21 pagesMetabolismo de Los CarbohidratosDANTESKO80No ratings yet
- Sistema EsqueleticoDocument23 pagesSistema Esqueleticoangel100% (3)
- Mitosis y MeiosisDocument21 pagesMitosis y MeiosisGabriela CuellarNo ratings yet
- Tejido ConectivoDocument41 pagesTejido ConectivoJuan Diego LosadaNo ratings yet
- LÍPIDOSDocument10 pagesLÍPIDOSRafael Angel Huiza Trujillo100% (3)
- Los Flagelos y CiliosDocument3 pagesLos Flagelos y CiliosMaia Roberts GFNo ratings yet
- La Reproducción CelularDocument4 pagesLa Reproducción CelularAlvaro100% (1)
- Tejidos EpitelialDocument45 pagesTejidos EpitelialLeslie Samantha Alvarado AguilarNo ratings yet
- Organelos CelularesDocument28 pagesOrganelos CelularesLogan Howlett100% (1)
- 2 2 La Membrana CelularDocument128 pages2 2 La Membrana Celulargraado100% (1)
- Tipos de CélulasDocument32 pagesTipos de CélulasCristina Isabel Maraboli Sandoval100% (1)
- Orgánulos No MembranososDocument5 pagesOrgánulos No Membranososalberto_fernandez_92100% (3)
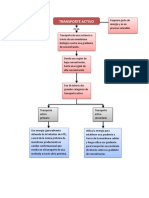
- Transporte CelularDocument10 pagesTransporte CelularAndres UrietaNo ratings yet
- 9 Tejido ConectivoDocument25 pages9 Tejido ConectivoJoel Fank Diaz Yopla100% (2)
- Tejidos Del Cuerpo HumanoDocument2 pagesTejidos Del Cuerpo Humanozapifiesta100% (1)
- Cavidad BucalDocument13 pagesCavidad BucalfelixabreuaNo ratings yet
- Generalidades en BacteriologíaDocument35 pagesGeneralidades en BacteriologíaBrenda Collantes VidalNo ratings yet
- Organelos de La Célula Animal y VegetalDocument3 pagesOrganelos de La Célula Animal y Vegetalruz marina garrido De Castillo100% (1)
- La CelulaDocument42 pagesLa Celulageysell arely100% (4)
- Aparato de GolgiDocument35 pagesAparato de GolgiJose Angel PerezNo ratings yet
- Cuadro Comparativo TejidosDocument3 pagesCuadro Comparativo TejidosGabriel Rodriguez100% (1)
- Sustancia Fundamental Amorfa y FormeDocument33 pagesSustancia Fundamental Amorfa y FormeJessica Ureño100% (3)
- CelulaDocument27 pagesCelulaFransis Gonçalves100% (2)
- Tejido ÓseoDocument6 pagesTejido ÓseoEdgar Feernando Martinez Sanchez100% (1)
- Fisiologia de La SangreDocument123 pagesFisiologia de La SangreBPMNo ratings yet
- ProteinasDocument16 pagesProteinasStephania Zambrano Guerra100% (1)
- Celula Eucariota y ProcariotaDocument19 pagesCelula Eucariota y ProcariotaVictor MagañaNo ratings yet
- La CelulaDocument29 pagesLa CelulaYanira Soto MedinaNo ratings yet
- Musculo EstriadoDocument19 pagesMusculo EstriadoLuis E. Hernández GuzmánNo ratings yet
- Tejido EpitelialDocument6 pagesTejido EpitelialSergio Dávalos75% (4)
- Histología Del Tejido EpitelialDocument8 pagesHistología Del Tejido EpitelialcecibelNo ratings yet
- Metabolismo CelularDocument61 pagesMetabolismo CelularAndres Monroy100% (1)
- Metabolismo Del GlucógenoDocument20 pagesMetabolismo Del Glucógenoqlawdya123No ratings yet
- Sistema DigestivoDocument23 pagesSistema DigestivoMelissa Paulino ZavaletaNo ratings yet
- Estructura y Función CelularDocument4 pagesEstructura y Función CelularCarolina OrtizNo ratings yet
- Tejido ConectivoDocument39 pagesTejido ConectivoAlexandra C DC50% (2)
- Ciclo CelularDocument45 pagesCiclo CelularAngel Bernuy100% (1)
- Celulas Del Sistema InmuneDocument14 pagesCelulas Del Sistema InmuneAntonio RodriguezNo ratings yet
- Sistema Membranoso - Exposicion Biologia - Grupo FDocument34 pagesSistema Membranoso - Exposicion Biologia - Grupo FAmbar OrtegaNo ratings yet
- AnabolismoDocument19 pagesAnabolismoCrystal BurrisNo ratings yet
- CitocinasDocument18 pagesCitocinasDave GC100% (1)
- Nucleo CelularDocument35 pagesNucleo CelularBiología Celular E Histología Médica100% (1)
- Sesion 3 Sintesis de ProteinasDocument41 pagesSesion 3 Sintesis de ProteinasAnonymous me3HTbZbh100% (2)
- Anabolismo y CatabolismoDocument14 pagesAnabolismo y Catabolismocelss100% (2)
- Imagenologia (Reydel Gonzalez)Document34 pagesImagenologia (Reydel Gonzalez)Reydel Gonzalez RodriguezNo ratings yet
- OvogénesisDocument80 pagesOvogénesisAlex100% (2)
- Tejido Epitelial - CaracterísticasDocument73 pagesTejido Epitelial - CaracterísticasMelany FrancescaNo ratings yet
- MetabolismoDocument15 pagesMetabolismoBryan Carrera100% (1)
- Resumen Capitulo Vi Economia Empresarial4Document23 pagesResumen Capitulo Vi Economia Empresarial4Misuzu KamioNo ratings yet
- Tabla Periodica 1 PDFDocument2 pagesTabla Periodica 1 PDFMisuzu KamioNo ratings yet
- Reporte Tecnología FarmacéuticaDocument11 pagesReporte Tecnología FarmacéuticaMisuzu KamioNo ratings yet
- Las 5 Supercomputadoras Más Potentes Del MundoDocument1 pageLas 5 Supercomputadoras Más Potentes Del MundoMisuzu KamioNo ratings yet
- ToxiDocument2 pagesToxiMisuzu KamioNo ratings yet
- Condiciones Grales Estandarizadas Final 21062016 PDFDocument25 pagesCondiciones Grales Estandarizadas Final 21062016 PDFMisuzu KamioNo ratings yet
- Puntos A Incluir en El ReporteDocument2 pagesPuntos A Incluir en El ReporteMisuzu KamioNo ratings yet
- Discusion Nutri Practica 2Document10 pagesDiscusion Nutri Practica 2Misuzu KamioNo ratings yet
- 05a PáncreasDocument51 pages05a PáncreasMisuzu KamioNo ratings yet
- Analisis Reporte 1Document2 pagesAnalisis Reporte 1Misuzu KamioNo ratings yet
- Previo 2Document2 pagesPrevio 2Misuzu KamioNo ratings yet
- Guia FarmaciasDocument16 pagesGuia FarmaciaskarmatrosNo ratings yet
- FormatoDocument4 pagesFormatoMisuzu KamioNo ratings yet
- Unidad de MezclasDocument8 pagesUnidad de MezclasMisuzu KamioNo ratings yet
- Previo 1 BiliotecaDocument12 pagesPrevio 1 BiliotecaMisuzu KamioNo ratings yet
- Anti Acid OsDocument4 pagesAnti Acid OsMisuzu KamioNo ratings yet
- Optativas y PaquetesLic en FarmaciaDocument21 pagesOptativas y PaquetesLic en FarmaciaMisuzu KamioNo ratings yet
- Resultados DiureticosDocument2 pagesResultados DiureticosMisuzu KamioNo ratings yet
- Introduccion Rep 2Document1 pageIntroduccion Rep 2Misuzu KamioNo ratings yet
- Plan de AccionDocument1 pagePlan de AccionMisuzu KamioNo ratings yet
- Trabajo Escrito de Interacciones FarmacologicasDocument10 pagesTrabajo Escrito de Interacciones FarmacologicasMisuzu KamioNo ratings yet
- Interacciones FarmacológicasDocument8 pagesInteracciones FarmacológicasMisuzu KamioNo ratings yet
- Trabajo Organica IndolDocument17 pagesTrabajo Organica IndolMisuzu KamioNo ratings yet
- Referencias Previo 2Document1 pageReferencias Previo 2Misuzu KamioNo ratings yet
- Difusión SimpleDocument4 pagesDifusión SimpleMisuzu KamioNo ratings yet
- En CabezadoDocument1 pageEn CabezadoMisuzu KamioNo ratings yet
- PosologiaDocument6 pagesPosologiaMisuzu KamioNo ratings yet
- BIOTERIODocument23 pagesBIOTERIODiego AlvaradoNo ratings yet
- Sustentacion Cariotipo y CromosomasDocument19 pagesSustentacion Cariotipo y CromosomasCAMILO ANDRES CORTINA ALFARONo ratings yet
- MeiosisDocument22 pagesMeiosisIris Betzaida Molina PintorNo ratings yet
- Promoción y Prevención Vacunación GeneralDocument39 pagesPromoción y Prevención Vacunación GeneralfannyNo ratings yet
- Cambios Sistemicos en El Paciente Adulto Mayor...Document3 pagesCambios Sistemicos en El Paciente Adulto Mayor...whysinpNo ratings yet
- Clonacion PresentacionDocument14 pagesClonacion PresentacionMaria CastilloNo ratings yet
- (Mitosis y Tarea)Document3 pages(Mitosis y Tarea)Carlos Antonio Mendoza ArteagaNo ratings yet
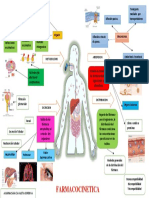
- Farmacocinetica - Mapa ConceptualDocument1 pageFarmacocinetica - Mapa Conceptualsara zavaleta cordova100% (1)
- PROF. Irma Cruzalegui Falcón - Alfonso Yauli Gutierrez Construimos Un País Libre de Discriminación Con Una Ciudadanía Crítica y ActivaDocument6 pagesPROF. Irma Cruzalegui Falcón - Alfonso Yauli Gutierrez Construimos Un País Libre de Discriminación Con Una Ciudadanía Crítica y ActivaLAHUANAMPA CHAVEZ AYDEE ROCIONo ratings yet
- Grado en Medicina 2012-2013 PDFDocument408 pagesGrado en Medicina 2012-2013 PDFCarmen Fernandez GarciaNo ratings yet
- Abstracts2010 PDFDocument193 pagesAbstracts2010 PDFriversgardenNo ratings yet
- TALLERDocument36 pagesTALLERMarianaNo ratings yet
- Ciclo CelularDocument5 pagesCiclo CelularEstradaNo ratings yet
- 2023 Valerani ADN Y NÚCLEO - 231004 - 110015Document26 pages2023 Valerani ADN Y NÚCLEO - 231004 - 110015Pedro PedroNo ratings yet
- Inhibidores de La ProteasaDocument2 pagesInhibidores de La ProteasaClaudia PeñaNo ratings yet
- Aterosclerosis IDocument1 pageAterosclerosis IMarco Antonio T. JiménezNo ratings yet
- LÍPIDOS TareaDocument6 pagesLÍPIDOS TareaKatrielManuelValladolidCastilloNo ratings yet
- Biotecnologia ForenseDocument4 pagesBiotecnologia Forensedynamo towNo ratings yet
- Tinción y Observación de MicroorganismosDocument4 pagesTinción y Observación de MicroorganismosJovenarellanoNo ratings yet
- Organizador Gráfico - Rutas Metabólicas Aerobias y AnaerobiasDocument1 pageOrganizador Gráfico - Rutas Metabólicas Aerobias y AnaerobiasElizabeth GarcíaNo ratings yet
- CromosomaDocument4 pagesCromosomalorenyis maitaNo ratings yet
- Fase1 - Actividad Colaborativa - Organismos TransgenicosDocument25 pagesFase1 - Actividad Colaborativa - Organismos Transgenicosvaness100% (1)
- Para Los Investigadores de Las Ciencias de La VidaDocument4 pagesPara Los Investigadores de Las Ciencias de La Vidasam taNo ratings yet
- BLADDocument10 pagesBLADNatalia GonzálezNo ratings yet
- Grado NovenoDocument26 pagesGrado Novenoayacucho01No ratings yet
- Cómo Se Produce La Fecundación HumanaDocument4 pagesCómo Se Produce La Fecundación HumanaJohn UC Diaz0% (1)
- Discusion #5 Genetica en MedicinaDocument7 pagesDiscusion #5 Genetica en MedicinaJulissa Giselle Campos LópezNo ratings yet
- Richard Escalona Semana 3Document3 pagesRichard Escalona Semana 3Richard Escalona100% (1)
- Tríptico COVID Formato TripticoDocument2 pagesTríptico COVID Formato TripticoLizeth SaldivarNo ratings yet
- Célula AnimalDocument4 pagesCélula AnimalEli Martinez AlfaroNo ratings yet