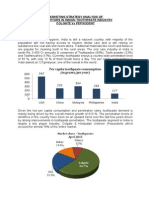
You might also like
- The Syn Alia Series On Animal TrainingDocument101 pagesThe Syn Alia Series On Animal Trainingben100% (1)
- Comparitive Study Colgate and PepsodentDocument10 pagesComparitive Study Colgate and PepsodentSunil MathewsNo ratings yet
- The Unified Neutral Theory of Biodiversity and Biogeography (MPB-32)From EverandThe Unified Neutral Theory of Biodiversity and Biogeography (MPB-32)Rating: 5 out of 5 stars5/5 (1)
- (Harrison & Cornell, 2008) Toward A Better Understanding of The Regional Causes of Local Community RichnessDocument11 pages(Harrison & Cornell, 2008) Toward A Better Understanding of The Regional Causes of Local Community RichnessOscar Leonardo Aaron Arizpe VicencioNo ratings yet
- Second Quarter Exam Pe 12Document4 pagesSecond Quarter Exam Pe 12Melegin Raton67% (3)
- Gem5 Splash2Document13 pagesGem5 Splash2Apurv SahayNo ratings yet
- (Ricklefs, 2008) Disintegration of Community Concept PDFDocument10 pages(Ricklefs, 2008) Disintegration of Community Concept PDFOscar Leonardo Aaron Arizpe VicencioNo ratings yet
- Does God Roll Dice? Neutrality and Determinism in Evolutionary EcologyDocument16 pagesDoes God Roll Dice? Neutrality and Determinism in Evolutionary EcologyAlara AğalarNo ratings yet
- Vellend Community Ecology QRB 2010Document25 pagesVellend Community Ecology QRB 2010liliana.piattiNo ratings yet
- Cavender Bares Etal2009Document23 pagesCavender Bares Etal2009Loana AraujoNo ratings yet
- 2007 Novack-Gottshall EcospaceDocument32 pages2007 Novack-Gottshall EcospaceobaytutNo ratings yet
- Vellend Community Ecology QRB 2010Document25 pagesVellend Community Ecology QRB 2010Calvince OdhiamboNo ratings yet
- Dynamics of Terrestrial GastropodsDocument24 pagesDynamics of Terrestrial GastropodsDerrick Scott FullerNo ratings yet
- From Populations to Ecosystems: Theoretical Foundations for a New Ecological Synthesis (MPB-46)From EverandFrom Populations to Ecosystems: Theoretical Foundations for a New Ecological Synthesis (MPB-46)No ratings yet
- Scale, Heterogeneity, and the Structure and Diversity of Ecological CommunitiesFrom EverandScale, Heterogeneity, and the Structure and Diversity of Ecological CommunitiesNo ratings yet
- Ecological Resilience, Biodiversity, and ScaleDocument13 pagesEcological Resilience, Biodiversity, and ScaleGerardo RiveroNo ratings yet
- Ecological Society of AmericaDocument14 pagesEcological Society of AmericaCleiton Lopes AguiarNo ratings yet
- Journal of Applied Ecology - 2012 - Johnson - Urbanization and Wetland Communities Applying Metacommunity Theory ToDocument9 pagesJournal of Applied Ecology - 2012 - Johnson - Urbanization and Wetland Communities Applying Metacommunity Theory ToSorayya Sangcopan OmarNo ratings yet
- How Do Biodiversity and Culture IntersectDocument17 pagesHow Do Biodiversity and Culture IntersectcaspianforestNo ratings yet
- 1269 MofddDocument9 pages1269 MofddMalosGodelNo ratings yet
- Grinnellian and Eltonian Niches and Geographic Distributions of SpeciesDocument9 pagesGrinnellian and Eltonian Niches and Geographic Distributions of SpeciesLeonardo CermeloNo ratings yet
- Frontiers of EcologyDocument10 pagesFrontiers of EcologylilujunNo ratings yet
- Post - Eco-Evolutionary Feedbacks in Community and Ecosystem Ecology (2009) 12pDocument12 pagesPost - Eco-Evolutionary Feedbacks in Community and Ecosystem Ecology (2009) 12pmartin686No ratings yet
- Landscape Ecology, Cross-Disciplinarity, and Sustainability ScienceDocument4 pagesLandscape Ecology, Cross-Disciplinarity, and Sustainability ScienceThinh NguyenNo ratings yet
- Community Ecology: Learning OutcomesDocument16 pagesCommunity Ecology: Learning OutcomesJezza Mae Gomba RegidorNo ratings yet
- Functional Diversity of Crustacean Zooplankton Communities Towards A Trait-Based Classification 2007 BARNETT Marcado PdfviewDocument18 pagesFunctional Diversity of Crustacean Zooplankton Communities Towards A Trait-Based Classification 2007 BARNETT Marcado PdfviewrgelabertNo ratings yet
- Trophic Theory of Island BiogeographyDocument8 pagesTrophic Theory of Island BiogeographyAlejandro Guerrero LaverdeNo ratings yet
- Ecological Concepts Involving Niche TheoryDocument7 pagesEcological Concepts Involving Niche TheoryNdutu MunywokiNo ratings yet
- Human+Ecology+and+its+applications LawrenceDocument10 pagesHuman+Ecology+and+its+applications LawrencelaxmanabcdNo ratings yet
- Ohlmann Et Al 2019 Div NetworksDocument33 pagesOhlmann Et Al 2019 Div Networkskacagj bajazzoNo ratings yet
- Balee 2006 - The Research Program of Historical EcologyDocument24 pagesBalee 2006 - The Research Program of Historical EcologyJuan VillanuevaNo ratings yet
- Historical Biogeography, Ecology and Species Richness: John J. Wiens and Michael J. DonoghueDocument6 pagesHistorical Biogeography, Ecology and Species Richness: John J. Wiens and Michael J. DonoghuePatricia Matú ÁlvarezNo ratings yet
- Ecological Thinking For BiodiversityDocument16 pagesEcological Thinking For BiodiversityAin NajwaNo ratings yet
- Conservation Biology and Traditional Ecological Knowledge: Integrating Academic Disciplines For Better Conservation PracticeDocument9 pagesConservation Biology and Traditional Ecological Knowledge: Integrating Academic Disciplines For Better Conservation PracticeSombra del CronopioNo ratings yet
- 2020 - Lagomarsino & Frost - The Central Role of Taxonomy in The Study of Neotropical BiodeiversityDocument17 pages2020 - Lagomarsino & Frost - The Central Role of Taxonomy in The Study of Neotropical BiodeiversityArturoCosmasAvilaNo ratings yet
- 1.0 Coevolutionary Ecological EconomicsDocument10 pages1.0 Coevolutionary Ecological Economicspuyehue123No ratings yet
- A Link Between Plant Traits and Abundance: Evidence From Coastal California Woody PlantsDocument8 pagesA Link Between Plant Traits and Abundance: Evidence From Coastal California Woody PlantsLigia Paulillo SimsNo ratings yet
- Ecological Succession in A Changing World: Cynthia C. Chang - Benjamin L. TurnerDocument7 pagesEcological Succession in A Changing World: Cynthia C. Chang - Benjamin L. TurnerRismatul JannahNo ratings yet
- Biotic Homogenization and Changes in Species Diversity Across Human Modified EcosystemsDocument8 pagesBiotic Homogenization and Changes in Species Diversity Across Human Modified Ecosystemsmp2233No ratings yet
- Andresen Etal 2018 Chapter1 BiodiversityDocument14 pagesAndresen Etal 2018 Chapter1 BiodiversityShubham SahaNo ratings yet
- Landscape EcologyDocument19 pagesLandscape Ecologyargie joy marieNo ratings yet
- Soil Fauna and Soil FunctionsDocument37 pagesSoil Fauna and Soil FunctionsAkmalNo ratings yet
- N ICHE C ONSERVATISM: Integrating Evolution, Ecology, and Conservation BiologyDocument23 pagesN ICHE C ONSERVATISM: Integrating Evolution, Ecology, and Conservation BiologyLaura Teresa PinzónNo ratings yet
- Interactions Among Plants and Evolution: Essay ReviewDocument12 pagesInteractions Among Plants and Evolution: Essay Reviewanon_456200170No ratings yet
- Baleé-14-Historical Ecology and The Explanation of Diversity-Amazonian Case StudiesDocument15 pagesBaleé-14-Historical Ecology and The Explanation of Diversity-Amazonian Case StudiesPaoliitha GuerreroNo ratings yet
- Ellis 2015 (Ecology)Document45 pagesEllis 2015 (Ecology)alianzacampeonNo ratings yet
- Redman 2005 Resilence Theory in ArchaeologyDocument8 pagesRedman 2005 Resilence Theory in ArchaeologygiustamarcoNo ratings yet
- Ecology (from Greek: οDocument40 pagesEcology (from Greek: οAnonymous E4Rbo2sNo ratings yet
- Walkup Et Al. (2017)Document15 pagesWalkup Et Al. (2017)Ruben FernandezNo ratings yet
- McCann The Diversity-Stability DebateDocument7 pagesMcCann The Diversity-Stability DebateFernando DuicaNo ratings yet
- 1.abundance & DiversityDocument170 pages1.abundance & DiversityFaisal JavidNo ratings yet
- CH 2 Fundamental Concept of EcologyDocument12 pagesCH 2 Fundamental Concept of EcologyyayaNo ratings yet
- Applying Predictive Models To Study The Ecological Proper - 2021 - Landscape andDocument13 pagesApplying Predictive Models To Study The Ecological Proper - 2021 - Landscape andImagine boyNo ratings yet
- Does Diversity Equal Stability? Are Diverse Ecosystems The Most Stable Ecosystems?Document2 pagesDoes Diversity Equal Stability? Are Diverse Ecosystems The Most Stable Ecosystems?LaurenSwannNo ratings yet
- Biodiversity and Environmental StabilityDocument10 pagesBiodiversity and Environmental StabilityGreenthingsNo ratings yet
- Mcart Et Al. (2011)Document9 pagesMcart Et Al. (2011)robertoNo ratings yet
- Fractal NatureDocument8 pagesFractal NatureComunidade do ReinoNo ratings yet
- Musa Cchio 2011Document5 pagesMusa Cchio 2011Benjan't PujadasNo ratings yet
- Webb Et Al 2002 Phylogenies and Community EcologyDocument32 pagesWebb Et Al 2002 Phylogenies and Community EcologyDevlynn CoelhoNo ratings yet
- CH 3 Rozzi & Tauro Field Environmental Philosophy Springer 2023Document25 pagesCH 3 Rozzi & Tauro Field Environmental Philosophy Springer 2023Enzo Navarrete UlloaNo ratings yet
- Ecosystem Responses To Community DisassemblyDocument23 pagesEcosystem Responses To Community DisassemblyDiana MariaNo ratings yet
- A Taxonomic Wish-List For Community Ecology: Nicholas J. GotelliDocument13 pagesA Taxonomic Wish-List For Community Ecology: Nicholas J. GotelliMarlon Cezar CominettiNo ratings yet
- Human Natures, Nature Conservation, and Environmental EthicsDocument13 pagesHuman Natures, Nature Conservation, and Environmental EthicsRowaci IdNo ratings yet
- Beatriz Milhazes Interview: There Is Nothing Simple About What I'm Doing'Document2 pagesBeatriz Milhazes Interview: There Is Nothing Simple About What I'm Doing'C_RigbyNo ratings yet
- Knowledge For You (Knowledge4u.org) Magazine Volume 1 Issue 1Document4 pagesKnowledge For You (Knowledge4u.org) Magazine Volume 1 Issue 1Brîndușa PetruțescuNo ratings yet
- What Is A Separate Legal Entity of A CompanyDocument7 pagesWhat Is A Separate Legal Entity of A CompanyTalha shabirNo ratings yet
- The Myth of The Indian Middle Class TEXTDocument2 pagesThe Myth of The Indian Middle Class TEXTSumaiya Shaheedha AnvarNo ratings yet
- Surface Tension Lab - ProcedureDocument3 pagesSurface Tension Lab - Procedurekatherine corveraNo ratings yet
- ZONE A - Walled City ReportDocument49 pagesZONE A - Walled City ReportAr Vishnu Prakash0% (1)
- Technical Information: Spare Eebds Mentioned in Item 1 Are Not To Be Included in Total NumberDocument5 pagesTechnical Information: Spare Eebds Mentioned in Item 1 Are Not To Be Included in Total NumberSatbir SinghNo ratings yet
- Equipment List ElectricalDocument4 pagesEquipment List ElectricalKaustabha DasNo ratings yet
- Lim Tanhu v. Ramolete (G.R. No. L-40098 August 29, 1975)Document48 pagesLim Tanhu v. Ramolete (G.R. No. L-40098 August 29, 1975)Hershey Delos SantosNo ratings yet
- Read The Text and Fill The Gaps With The Words Given BelowDocument2 pagesRead The Text and Fill The Gaps With The Words Given Belownatomasurashvili3100% (1)
- Toad For Oracle Release Notes 2017 r2Document30 pagesToad For Oracle Release Notes 2017 r2Plate MealsNo ratings yet
- 7-8 Seminary PresentationDocument8 pages7-8 Seminary PresentationDumitru Ioana RuxandraNo ratings yet
- Chap 3Document11 pagesChap 3Quỳnh GiaoNo ratings yet
- List of Telugu Films of 2005 - WikipediaDocument1 pageList of Telugu Films of 2005 - WikipediaRa RsNo ratings yet
- 8-31 AhlamDocument14 pages8-31 AhlamJeff BowmanNo ratings yet
- SOAL UTS Poetry Genap 2020Document5 pagesSOAL UTS Poetry Genap 2020Zara AzkiyahNo ratings yet
- ELC102 - Episode 1Document3 pagesELC102 - Episode 1Juzwa MaramotNo ratings yet
- Affective Evaluation: Group 5: 1. Yana Sofia Putri 2. Mamanda Silitonga 3. Sarah Nur Fadhilah PanjaitanDocument36 pagesAffective Evaluation: Group 5: 1. Yana Sofia Putri 2. Mamanda Silitonga 3. Sarah Nur Fadhilah PanjaitanGresia FalentinaNo ratings yet
- Introduction of TECHNIP PDFDocument123 pagesIntroduction of TECHNIP PDFdigeca100% (1)
- EmpBen TheoriesDocument8 pagesEmpBen TheoriesCarl Dhaniel Garcia SalenNo ratings yet
- On JioDocument10 pagesOn JioSimranNo ratings yet
- United States v. Ivan Nikonorovich Rogalsky, 575 F.2d 457, 3rd Cir. (1978)Document6 pagesUnited States v. Ivan Nikonorovich Rogalsky, 575 F.2d 457, 3rd Cir. (1978)Scribd Government DocsNo ratings yet
- Power Plant Economics 1 (UNIT-I)Document21 pagesPower Plant Economics 1 (UNIT-I)rahul soniNo ratings yet
- Socio Cultural Factors Influencing Child Nutrition Among Mothers in Calabar Municipality Cross River State NigeriaDocument34 pagesSocio Cultural Factors Influencing Child Nutrition Among Mothers in Calabar Municipality Cross River State NigeriaTAJUDEEN AZEEZNo ratings yet
- Animal Farm: Social and Historical ContextDocument4 pagesAnimal Farm: Social and Historical ContextfancyNo ratings yet
- 13 - Chapter 3Document22 pages13 - Chapter 3manoj varmaNo ratings yet