You might also like
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Hyperalgesia in The Setting of AnxietyDocument1 pageHyperalgesia in The Setting of AnxietynikuNo ratings yet
- Curriculum Vitae: ExperienceDocument2 pagesCurriculum Vitae: ExperiencenikuNo ratings yet
- Sunita Apticon15Document3 pagesSunita Apticon15nikuNo ratings yet
- Scott Reuschel TANDEM LABS - Regulated Bioanlysis For The Pharmaceutical IndustryDocument33 pagesScott Reuschel TANDEM LABS - Regulated Bioanlysis For The Pharmaceutical IndustrynikuNo ratings yet
- Following OvulationDocument1 pageFollowing OvulationnikuNo ratings yet
- Balram Jadhav CVDocument3 pagesBalram Jadhav CVnikuNo ratings yet
- Synopsis EditedDocument8 pagesSynopsis EditednikilNo ratings yet
- PKCDocument52 pagesPKCnikuNo ratings yet
- Genome Mapping: Paul H DearDocument7 pagesGenome Mapping: Paul H DearvidpoohNo ratings yet
- CytokininDocument7 pagesCytokininPablo AlbanNo ratings yet
- Overdose Dan ToxikasiDocument44 pagesOverdose Dan ToxikasiRandi N. FarizaNo ratings yet
- ELSI Working Group 1st ReportDocument9 pagesELSI Working Group 1st ReportnikuNo ratings yet
- NeuropeptidesDocument36 pagesNeuropeptidesnikuNo ratings yet
- Chrono 3Document9 pagesChrono 3nikuNo ratings yet
- Systemic Target Single ExposureDocument10 pagesSystemic Target Single ExposurenikuNo ratings yet
- Chrono 4Document27 pagesChrono 4nikuNo ratings yet
- Management PoisoningDocument6 pagesManagement PoisoningatifdocNo ratings yet
- Pharmacologic Activity of Phosphodiesterases and Their InhibitorsDocument15 pagesPharmacologic Activity of Phosphodiesterases and Their InhibitorsnikuNo ratings yet
- Hyperalgesia in The Setting of AnxietyDocument1 pageHyperalgesia in The Setting of AnxietynikuNo ratings yet
- Chrono 2Document5 pagesChrono 2nikuNo ratings yet
- Teach Tox SlidesDocument23 pagesTeach Tox SlidesnikuNo ratings yet
- Systemin Target Repeated ExposureDocument11 pagesSystemin Target Repeated ExposurenikuNo ratings yet
- Pro Kinase CDocument9 pagesPro Kinase CnikuNo ratings yet
- Week2PostHoc 03Document22 pagesWeek2PostHoc 03nikuNo ratings yet
- United States: (12) Patent Application Publication (10) Pub. No.: US 2013/0052239 A1Document22 pagesUnited States: (12) Patent Application Publication (10) Pub. No.: US 2013/0052239 A1nikuNo ratings yet
- NIH Public Access: Author ManuscriptDocument24 pagesNIH Public Access: Author ManuscriptnikuNo ratings yet
- United States: (12) Patent Application Publication (10) Pub. No.: US 2011/0206766 A1Document20 pagesUnited States: (12) Patent Application Publication (10) Pub. No.: US 2011/0206766 A1nikuNo ratings yet
- Us 20100323011Document15 pagesUs 20100323011nikuNo ratings yet
- NPP 2009222 ADocument12 pagesNPP 2009222 AnikuNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Retinoid Receptors and Therapeutic Applications of RAR/RXR ModulatorsDocument23 pagesRetinoid Receptors and Therapeutic Applications of RAR/RXR ModulatorsAKellyNMNo ratings yet
- 155:411 Introduction To Biochemical Engineering Fall 2012 Exam 1 October 10, 2012 From 5:00-8:00 P.MDocument2 pages155:411 Introduction To Biochemical Engineering Fall 2012 Exam 1 October 10, 2012 From 5:00-8:00 P.Mmanthan2120% (1)
- Confidential 1 Pr110 WatermarkDocument51 pagesConfidential 1 Pr110 WatermarkMarissa AldeNo ratings yet
- Nutrition Answers FinalDocument9 pagesNutrition Answers FinalelizabethNo ratings yet
- DLP - Tomatom. Science 10. q3 - DNA REPLICAtionDocument10 pagesDLP - Tomatom. Science 10. q3 - DNA REPLICAtionalchristian tomatomNo ratings yet
- MS MLPA Protocol One Tube MSP v008Document19 pagesMS MLPA Protocol One Tube MSP v008Sp PpvNo ratings yet
- Evolution-History and EvidenceDocument19 pagesEvolution-History and Evidencekatexxx100% (1)
- General Biology 1 Summative Assessment Week 1-4Document2 pagesGeneral Biology 1 Summative Assessment Week 1-4Cherrina Aguila100% (1)
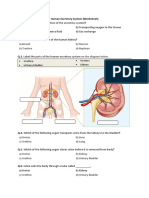
- Human Excretory SystemDocument2 pagesHuman Excretory SystemVio ElenaNo ratings yet
- Lec 2 Part 2Document34 pagesLec 2 Part 2Ahmed JamalNo ratings yet
- Cytogenetics Transes Topic 16Document5 pagesCytogenetics Transes Topic 16Dianne NolascoNo ratings yet
- Sysoeva Chemical Composition Andbiological Activity of Triterpenes and Steroids of Chaga MushroomDocument8 pagesSysoeva Chemical Composition Andbiological Activity of Triterpenes and Steroids of Chaga MushroomMachfud FauziNo ratings yet
- Lecture 7Document68 pagesLecture 7Gay TonyNo ratings yet
- Usman Public School System: 1 Monthly Test Mid-Term 2018 - 2019Document3 pagesUsman Public School System: 1 Monthly Test Mid-Term 2018 - 2019Huzaifa KhanNo ratings yet
- OPfr4 2Document13 pagesOPfr4 26E Rivera Cervantes Osiris MargaritaNo ratings yet
- J Zool Syst Evol Res - 2020 - Khan - Molecular Phylogeny of Eremias SPP From Pakistan Contributes To A BetterDocument18 pagesJ Zool Syst Evol Res - 2020 - Khan - Molecular Phylogeny of Eremias SPP From Pakistan Contributes To A BettershakeeNo ratings yet
- Daftar Pustaka PembahasanDocument2 pagesDaftar Pustaka PembahasanMaryam Fathiya RinthaniNo ratings yet
- Green Life Cycle Science Butterfly Graphic OrganizerDocument1 pageGreen Life Cycle Science Butterfly Graphic OrganizerAlexia SalvadorNo ratings yet
- Rocafort Et Al., 2020 Apoplastic Effector Proteins of Plant-Associated Fungi and Oomycetes PDFDocument11 pagesRocafort Et Al., 2020 Apoplastic Effector Proteins of Plant-Associated Fungi and Oomycetes PDFJuan Muñoz CuellarNo ratings yet
- Week 2 - Text 1. Ecosystems and Human InterferencesDocument2 pagesWeek 2 - Text 1. Ecosystems and Human Interferences458 269 Cinta Alivia AthaNo ratings yet
- Biology Navneet Practice Paper - 240102 - 221900Document27 pagesBiology Navneet Practice Paper - 240102 - 221900deepak hutthyNo ratings yet
- Clinical Radio Biology - Previous Examination Papers Updated 040108Document12 pagesClinical Radio Biology - Previous Examination Papers Updated 040108sawiHijau100% (3)
- Anatomi TumbuhanDocument30 pagesAnatomi TumbuhanMEISYI DSNo ratings yet
- TuberculosisDocument16 pagesTuberculosisjojolim18100% (1)
- HSA. Past Papers With Answer SDocument33 pagesHSA. Past Papers With Answer SAaron Baker100% (1)
- Els Reviewer 2nd QuarterDocument8 pagesEls Reviewer 2nd QuarterKaezzy Yen RequermeNo ratings yet
- Acinetobacter BaumanniiDocument12 pagesAcinetobacter BaumanniiGIST (Gujarat Institute of Science & Technology)No ratings yet
- XXXDocument134 pagesXXXrhlzuraNo ratings yet
- Ijms 23 03754Document14 pagesIjms 23 03754Benyam ZenebeNo ratings yet
- CHAPTER 16 MicroparaDocument46 pagesCHAPTER 16 MicroparaKathlyn LopeñaNo ratings yet