You might also like
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Coimbatore Corporation Master Plan Map 2021Document1 pageCoimbatore Corporation Master Plan Map 2021Rahul KumarNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Usage of NCL, Grads, Pyhdf, GDL and Gdal To Access HDF FilesDocument39 pagesUsage of NCL, Grads, Pyhdf, GDL and Gdal To Access HDF FilesRahul KumarNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- Determination of Iron by Atomic Absorption SpectrophotometryDocument3 pagesDetermination of Iron by Atomic Absorption SpectrophotometryRahul KumarNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- HDF-EOS5 Data Model, File Format and LibraryDocument56 pagesHDF-EOS5 Data Model, File Format and LibraryRahul KumarNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Pipetting Guide - Thermo Scientific - 25440 PDFDocument20 pagesPipetting Guide - Thermo Scientific - 25440 PDFRahul KumarNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Adaptive ImmunityDocument25 pagesAdaptive ImmunityRahul KumarNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Population Genetics ProblemsDocument4 pagesPopulation Genetics ProblemsRahul KumarNo ratings yet
- Biodiesel From Algae: Challenges and ProspectsDocument10 pagesBiodiesel From Algae: Challenges and ProspectsRahul KumarNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The Downfall of The German Shepherd: by Koos HassingDocument15 pagesThe Downfall of The German Shepherd: by Koos HassingMustafa Ehlizevak100% (1)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Guia02 PDFDocument191 pagesGuia02 PDFdanymoloNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Facts About Respiratory SystemDocument10 pagesFacts About Respiratory SystemGermie Mariel GojarNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Summary Conclusion and RecommendationsDocument6 pagesSummary Conclusion and RecommendationsjmpangeloNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Jose R. Reyes Memorial Medical Center Department of Surgery: Clinical DatabaseDocument2 pagesJose R. Reyes Memorial Medical Center Department of Surgery: Clinical DatabaseMary Rose DomalantaNo ratings yet
- CLASSIC ARTICLE Clinical Measurement and EvaluationDocument5 pagesCLASSIC ARTICLE Clinical Measurement and EvaluationJesusCordoba100% (2)
- Tumori PancreasDocument71 pagesTumori PancreasAnda Madalina ZahariaNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
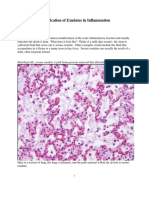
- Identification of Exudates in Inflammation...Document5 pagesIdentification of Exudates in Inflammation...Ivana Aginta GintingNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Insects and Spiders - Grzimek's SalrDocument531 pagesInsects and Spiders - Grzimek's SalrPoncho Efren De La CruzNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Latuske P, Et Al - Front Pharmacol. 2022 Oct 14 13898548.Document14 pagesLatuske P, Et Al - Front Pharmacol. 2022 Oct 14 13898548.刘林玉No ratings yet
- General Properties of VirusesDocument24 pagesGeneral Properties of VirusesPeachy PieNo ratings yet
- THE INTEGRATIVE ACTION OF THE NERVOUS SYSTEM - Charles SherringtonDocument442 pagesTHE INTEGRATIVE ACTION OF THE NERVOUS SYSTEM - Charles SherringtonGastón Tadeo ReyeroNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Hema 1A Lec 1 Shift Reviewer Hematopoiesis Prepared By: C Hematopoiesis DefinedDocument7 pagesHema 1A Lec 1 Shift Reviewer Hematopoiesis Prepared By: C Hematopoiesis DefinedMarc ViduyaNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Nasogastric+Tube+Insertion+and+Removal 2Document24 pagesNasogastric+Tube+Insertion+and+Removal 2Anonymous 4OJslWE1No ratings yet
- Essene FastingDocument48 pagesEssene FastingEnoch Gandhislilbrother Abraham90% (10)
- Cell Structure and OrganizationDocument18 pagesCell Structure and OrganizationMuhammad SamhanNo ratings yet
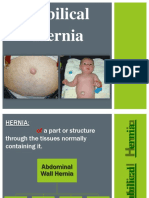
- Hernia UmbilikalisDocument16 pagesHernia UmbilikalisWibhuti EmrikoNo ratings yet
- Indirect Laryngeoscopy - ReflectionDocument3 pagesIndirect Laryngeoscopy - ReflectionDanica LayagNo ratings yet
- Chapter Two Biological Basis of Behavior: 2.1. Definition and General RemarkDocument5 pagesChapter Two Biological Basis of Behavior: 2.1. Definition and General RemarkArvella AlbayNo ratings yet
- Handp TemplateDocument3 pagesHandp TemplateLauren GrandpreNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Guruji 2Document176 pagesGuruji 2Cine Dada67% (6)
- WormsDocument20 pagesWormsAdebisi Jeleel AdekunleNo ratings yet
- Expansion and Characterization of Human Natural Killer (NK) CellsDocument5 pagesExpansion and Characterization of Human Natural Killer (NK) CellsManoj Kumar pandreNo ratings yet
- ROSE Chapter 2Document19 pagesROSE Chapter 2Gabriel PascualNo ratings yet
- MAPPING Ginekologi Onkologi 22-05-2018Document7 pagesMAPPING Ginekologi Onkologi 22-05-2018adityoNo ratings yet
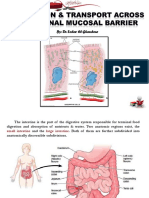
- Intestinal Mucosal Barrier & Highly Absorptive CellsDocument25 pagesIntestinal Mucosal Barrier & Highly Absorptive Cellsas qw100% (1)
- Pediatric Assessment Triangle EditDocument31 pagesPediatric Assessment Triangle EditAlbert Gunawan100% (1)
- Prevalence of Sarcoptic Mange in Rabbits: A. Meenakshisundaram and T. AnnaDocument6 pagesPrevalence of Sarcoptic Mange in Rabbits: A. Meenakshisundaram and T. AnnaAchmad NugrohoNo ratings yet
- Adjectives Starting With AlphabetsDocument647 pagesAdjectives Starting With AlphabetsHemanta majhiNo ratings yet
- Vet School Personal StatementDocument2 pagesVet School Personal Statementfinishstrong075756100% (1)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)