You might also like
- 1 Biología CelularDocument38 pages1 Biología CelularBRAYNER LOPEZ SARMIENTONo ratings yet
- Tema 7Document7 pagesTema 7Claudia Perujo VallejoNo ratings yet
- Estructura y Fisiología CelularDocument23 pagesEstructura y Fisiología CelularValentina PeraltaNo ratings yet
- Tema 7Document6 pagesTema 7Claudia Perujo VallejoNo ratings yet
- CelulasDocument58 pagesCelulasluz claraNo ratings yet
- Estructurasyorganelosdelacelula 10 Bio PDFDocument7 pagesEstructurasyorganelosdelacelula 10 Bio PDFPablo HernàndezNo ratings yet
- Esquema - Dibujo - Trabajo Practico Biologia CelularDocument4 pagesEsquema - Dibujo - Trabajo Practico Biologia CelulargerovixNo ratings yet
- Cel Pro EucDocument5 pagesCel Pro EucMauro Miguel Marsiglia MejiaNo ratings yet
- MICROBIOLOGIA GENERAL ResumenDocument90 pagesMICROBIOLOGIA GENERAL ResumenOriana MuruaNo ratings yet
- Funcion Celular, SeñalizaciónDocument63 pagesFuncion Celular, SeñalizaciónDaryl TinjacaNo ratings yet
- 3 Estructura Celular y Crecimiento de Los MicroorganismosDocument56 pages3 Estructura Celular y Crecimiento de Los MicroorganismosKenner Andrés Ayala NavarroNo ratings yet
- Unidad I-1-1Document31 pagesUnidad I-1-1Katerin VenturaNo ratings yet
- BiologiaDocument2 pagesBiologiaMaru UxiNo ratings yet
- Clase de La CélulaDocument29 pagesClase de La Célulamariolis espinozaNo ratings yet
- APUNTES SEMINARIOS BiologiaDocument103 pagesAPUNTES SEMINARIOS BiologiaborellaazulNo ratings yet
- La Estructura Celular Procariota Básica Tiene Los Siguientes ComponentesDocument6 pagesLa Estructura Celular Procariota Básica Tiene Los Siguientes ComponentesVictor CidNo ratings yet
- 2.1. La Celula - 2022-23Document65 pages2.1. La Celula - 2022-23sfdfsdfNo ratings yet
- Tema 8Document9 pagesTema 8Serena AchartNo ratings yet
- ¿Qué Son Los Ácidos Nucleicos?: EstructuraDocument8 pages¿Qué Son Los Ácidos Nucleicos?: EstructuraKaren Adriana Rangel Ramírez 1121No ratings yet
- Diplomado Morfologia Celular Hector Morante PDFDocument27 pagesDiplomado Morfologia Celular Hector Morante PDFRenzo BassanNo ratings yet
- Material de Estructura CelularDocument8 pagesMaterial de Estructura CelularKarina TorresNo ratings yet
- PAO 2 S3 DIAPOSITIVAS BIOCEL Semana 3Document25 pagesPAO 2 S3 DIAPOSITIVAS BIOCEL Semana 3Erika Geovanna Sinchi CarchichablaNo ratings yet
- QuimicaDocument5 pagesQuimicaTatiana Diaz CardonaNo ratings yet
- 5) Celulas - Microbianas - EucariotasDocument32 pages5) Celulas - Microbianas - EucariotasJulian BarreraNo ratings yet
- Célula - Fisica Médica 2do Semestre Medicina Udo AnzDocument19 pagesCélula - Fisica Médica 2do Semestre Medicina Udo AnzeduardovaleraNo ratings yet
- Clase 4 Componentes Quimicos de Las CelulasDocument69 pagesClase 4 Componentes Quimicos de Las Celulasraulquispecordova100% (1)
- Bloque Ii - BiologíaDocument12 pagesBloque Ii - Biologíafx9vjn75p8No ratings yet
- Citologia VegetalDocument7 pagesCitologia VegetalkathehernandezNo ratings yet
- Tema Celula Acidos Nucleicos-QcaOrgyBiol. - FI-FUNER-1C 2020Document46 pagesTema Celula Acidos Nucleicos-QcaOrgyBiol. - FI-FUNER-1C 2020Luciano RodríguezNo ratings yet
- 3 Cuadernillo. Características de Las CélulasDocument7 pages3 Cuadernillo. Características de Las Célulasa353862No ratings yet
- Celula TareaDocument3 pagesCelula TareaMauricio EduardoNo ratings yet
- Tema 4 Organizacion Del CitoplasmaDocument128 pagesTema 4 Organizacion Del CitoplasmaMinimo TNo ratings yet
- 3 - Células EucariotasDocument65 pages3 - Células EucariotasCris AcevedoNo ratings yet
- Cuadro Comparativo Celula Eucariota y ProcariotaDocument1 pageCuadro Comparativo Celula Eucariota y ProcariotaMaría José MarquezNo ratings yet
- Historia de La Biologia Celular @nit@Document13 pagesHistoria de La Biologia Celular @nit@anita_091951No ratings yet
- Citoplasma CelularDocument19 pagesCitoplasma CelularAlisson Caicedo100% (2)
- Microbiología Merged CompressedDocument147 pagesMicrobiología Merged CompressedNAYELI MELISA VASQUEZ CORREANo ratings yet
- MicrobiologíaDocument29 pagesMicrobiologíaEnrique Vera SagasteguiNo ratings yet
- LA CELULA Eucariota y ProcariotaDocument11 pagesLA CELULA Eucariota y Procariotaomegaxz7No ratings yet
- Diferencias de CelulasDocument3 pagesDiferencias de CelulasMartinez Gudiño ArturoNo ratings yet
- Celula Eucariota - Procariota - Ciclo CelularDocument9 pagesCelula Eucariota - Procariota - Ciclo CelularMARTINA CARROMBANo ratings yet
- Biologia GuiaDocument80 pagesBiologia GuiaalejandraNo ratings yet
- Equipo 3 - Caracteristicas Celulas Procariontas y EucariontasDocument21 pagesEquipo 3 - Caracteristicas Celulas Procariontas y EucariontasMiguel Angel Ruiz GuadarramaNo ratings yet
- Celulas Eucariotas y ProcariotasDocument57 pagesCelulas Eucariotas y Procariotaswalter100% (1)
- Celula Eucariota y ProcariotaDocument8 pagesCelula Eucariota y ProcariotaYurley PeñalozaNo ratings yet
- Cuadro Biologia BenjaDocument3 pagesCuadro Biologia BenjaTamara TrescherNo ratings yet
- Sesión 04 - Biología CelularDocument40 pagesSesión 04 - Biología CelularSILVINA JUDITH CABRERA MATTOSNo ratings yet
- Resumenes BiocelDocument25 pagesResumenes BiocelAna Kareli FríasNo ratings yet
- Resumen IBCM-1Document34 pagesResumen IBCM-1Micaela CoriaNo ratings yet
- Tema 3 PDFDocument23 pagesTema 3 PDFWanda DuarteNo ratings yet
- Biologia Guia N°2 8°basicoDocument12 pagesBiologia Guia N°2 8°basicoMARIA DEL CARMEN VIVANCONo ratings yet
- Cuadros para Ex - Biología EnfermeríaDocument14 pagesCuadros para Ex - Biología EnfermeríaF3n@nd0No ratings yet
- Histologia Resumen 497361 Downloable 2575799Document7 pagesHistologia Resumen 497361 Downloable 2575799Nicolás TorresNo ratings yet
- CLASE 3 - Eucariotas y Procariotas-VirusDocument25 pagesCLASE 3 - Eucariotas y Procariotas-Virusfiorellalzak1507No ratings yet
- N° 02 CelulaDocument53 pagesN° 02 CelulaYHUMY CARRION UTANINo ratings yet
- FISIOPATOLOGÍADocument4 pagesFISIOPATOLOGÍAMaría Grueso MuñozNo ratings yet
- Nucleo CelukarDocument3 pagesNucleo CelukarValeria MartinezNo ratings yet
- Cuestionario Sobre CromosomasDocument2 pagesCuestionario Sobre CromosomashernanNo ratings yet
- Clase 9 Microcuerpos, Plastidios, Mitocondrias VETER AGRONO 2020 20Document22 pagesClase 9 Microcuerpos, Plastidios, Mitocondrias VETER AGRONO 2020 20omar collasNo ratings yet
- Celula ProcariotaDocument3 pagesCelula ProcariotaAngel Fredy Tacza QuinteNo ratings yet
- Potenciales de Membrana y Potenciales de Accion PDFDocument53 pagesPotenciales de Membrana y Potenciales de Accion PDFPerla LP100% (13)
- 2 Prueba Guia Biologia Nivel 2M Mitosis y MeiosisDocument9 pages2 Prueba Guia Biologia Nivel 2M Mitosis y MeiosisNorth Cloths Paños del NorteNo ratings yet
- 2-4 ClasificaciónDocument23 pages2-4 ClasificaciónDiego Mariano Escamilla HernandezNo ratings yet
- Western Blot ABCAM PresentaciónDocument49 pagesWestern Blot ABCAM PresentaciónArmando GBNo ratings yet
- Resultado 1042468153 23032523468MOCO 0 0FIDocument4 pagesResultado 1042468153 23032523468MOCO 0 0FITobias ArizaNo ratings yet
- Laboratorio Bilogia Celular. Jesus Meriño.Document16 pagesLaboratorio Bilogia Celular. Jesus Meriño.Jesús MeriñoNo ratings yet
- Epitelios y GlandulasDocument76 pagesEpitelios y GlandulasJineth Ruiz RodriguezNo ratings yet
- Taller Componente CelularDocument2 pagesTaller Componente CelularAea Grondona0% (1)
- Western Blot 2020.2docxDocument6 pagesWestern Blot 2020.2docxYeiny Lisbeth Angarita ParalesNo ratings yet
- Trabajo MareDocument10 pagesTrabajo MareCristian Romero QuinteroNo ratings yet
- LISOSOMA PeroxmarielaDocument32 pagesLISOSOMA PeroxmarielaMariela HuertamsNo ratings yet
- Histología AnimalDocument7 pagesHistología AnimalErick OlayaNo ratings yet
- Cuestionario 1 Evaluación Parcial - Primer Cuestionario en LíneaDocument5 pagesCuestionario 1 Evaluación Parcial - Primer Cuestionario en LíneaAlessandro GuerrónNo ratings yet
- Primaria IDocument47 pagesPrimaria IEdinson SagumaNo ratings yet
- Efecto de Hormonas Sobre El Metabolismo de Las LípidosDocument4 pagesEfecto de Hormonas Sobre El Metabolismo de Las LípidosFanny Marlene Biersack0% (1)
- Ensayo Biología MolecularDocument2 pagesEnsayo Biología MolecularJuan Pablo Argel MuñozNo ratings yet
- Estado de Choque GeneralidadesDocument35 pagesEstado de Choque GeneralidadesPaola RubioNo ratings yet
- Actividad 3 Unidad 4 PDFDocument2 pagesActividad 3 Unidad 4 PDFFrida HernandezNo ratings yet
- Virus Del Papiloma HumanoDocument20 pagesVirus Del Papiloma HumanoHeisly LinarezNo ratings yet
- Resistencia BacterianaDocument25 pagesResistencia BacterianaLourdes Carvajal BenitezNo ratings yet
- Resumen Del Artículo Bases Moleculares de Las Acciones de La InsulinaDocument4 pagesResumen Del Artículo Bases Moleculares de Las Acciones de La InsulinaAldo AcostaNo ratings yet
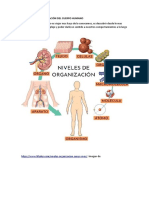
- Niveles y Organización Del Cuerpo HumanoDocument8 pagesNiveles y Organización Del Cuerpo HumanoMonica ManriqueNo ratings yet
- Actividad Integradora Proteínas - Relación Estructura-Función 2015Document2 pagesActividad Integradora Proteínas - Relación Estructura-Función 2015Silvia Mercedes BrogginiNo ratings yet
- Taller Herencia y GeneticaDocument6 pagesTaller Herencia y GeneticaVanessa VidarteNo ratings yet
- Evaluacion de LipidosDocument3 pagesEvaluacion de LipidosJeronimo OviedoNo ratings yet
- solINFORME DE LABORATORIO DE BIOLOGÍA PRACTICA RECONOCIMIENTO DE MATERIALESDocument9 pagessolINFORME DE LABORATORIO DE BIOLOGÍA PRACTICA RECONOCIMIENTO DE MATERIALESBELTRAN GALEANO JORGE ANTONIO100% (1)
- La Membrana Plasmática Regula La Entrada y Salida de MaterialesDocument2 pagesLa Membrana Plasmática Regula La Entrada y Salida de MaterialesDaicy Contreras DiazNo ratings yet
- BiomoleculasDocument11 pagesBiomoleculasTorres Reyes Josue AlbertoNo ratings yet