You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Latin Eugenics in Comparative PerspectiveDocument356 pagesLatin Eugenics in Comparative PerspectivemafeNo ratings yet
- ACLS Pretest Exam AnnotatedDocument21 pagesACLS Pretest Exam AnnotatedMOLLYNo ratings yet
- Media Violence Exposure and Physical Aggression in Fifth-Grade ChildrenDocument7 pagesMedia Violence Exposure and Physical Aggression in Fifth-Grade ChildrenAnonymous ceYk4p4No ratings yet
- Experiment Vii To Prepare A Temporary Mount of Onion Root Tip To Study MitosisDocument4 pagesExperiment Vii To Prepare A Temporary Mount of Onion Root Tip To Study MitosisAnonymous ceYk4p4100% (12)
- Acn NarangiDocument32 pagesAcn NarangiAnonymous ceYk4p4No ratings yet
- PV in India Good ArticleDocument8 pagesPV in India Good ArticleAnonymous ceYk4p4No ratings yet
- Application For Replacement of Faulty RefrigeratorDocument1 pageApplication For Replacement of Faulty RefrigeratorAnonymous ceYk4p4No ratings yet
- Boss Si Online FormDocument1 pageBoss Si Online FormAnonymous ceYk4p4No ratings yet
- List of Eligible Candidates and Rejected Applications For Interview For Short Term Paid Clinical Assistantship 2017Document2 pagesList of Eligible Candidates and Rejected Applications For Interview For Short Term Paid Clinical Assistantship 2017Anonymous ceYk4p4No ratings yet
- Application For Replacement of Faulty LPG RegulatorDocument1 pageApplication For Replacement of Faulty LPG RegulatorAnonymous ceYk4p40% (1)
- Instrs For Isg Detailed To Cover Prac CampDocument22 pagesInstrs For Isg Detailed To Cover Prac CampAnonymous ceYk4p4No ratings yet
- Entomology Question Paper 2005Document6 pagesEntomology Question Paper 2005Anonymous ceYk4p4100% (1)
- Advertisement For The Post of Dentist in Indian Air Force Station, DalhousieDocument1 pageAdvertisement For The Post of Dentist in Indian Air Force Station, DalhousieAnonymous ceYk4p4No ratings yet
- Post Insertion Complaints in CD For Dental Students For Final Year For Their Seminar.Document31 pagesPost Insertion Complaints in CD For Dental Students For Final Year For Their Seminar.Anonymous ceYk4p4No ratings yet
- Biomechanical Tooth Preparation Principles For BDS Final Year StudentsDocument80 pagesBiomechanical Tooth Preparation Principles For BDS Final Year StudentsAnonymous ceYk4p4100% (2)
- Call Letter List Dental - INDIAN ARMY DENTAL CORPS 2012Document24 pagesCall Letter List Dental - INDIAN ARMY DENTAL CORPS 2012Anonymous ceYk4p4No ratings yet
- BDS First Professional Exam Question Paper 2008-2009 - Teerthanker Mahaveer Dental CollegeDocument21 pagesBDS First Professional Exam Question Paper 2008-2009 - Teerthanker Mahaveer Dental CollegeAnonymous ceYk4p4No ratings yet
- Project ProposalDocument3 pagesProject ProposalSkrrtt SkrrttNo ratings yet
- Philo Final SummativeDocument2 pagesPhilo Final SummativeFobe Lpt NudaloNo ratings yet
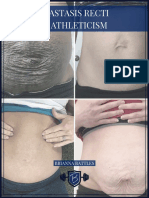
- Diastasis Recti Ebook DownloadDocument19 pagesDiastasis Recti Ebook DownloadMertcan DamatNo ratings yet
- 1833 TX Melton, Daisy Mae DeathDocument1 page1833 TX Melton, Daisy Mae DeathRichard TonsingNo ratings yet
- Nursing Care Plan: Phinma University of PangasinanDocument1 pageNursing Care Plan: Phinma University of PangasinanShaira De La CruzNo ratings yet
- Ufgs 01 57 19.01 20Document63 pagesUfgs 01 57 19.01 20jackcan501No ratings yet
- Clinic by Alissa QuartDocument4 pagesClinic by Alissa QuartOnPointRadioNo ratings yet
- Aj Infratech: Job Hazard AnalysisDocument5 pagesAj Infratech: Job Hazard AnalysisDwitikrushna RoutNo ratings yet
- The Pale Blood HuntDocument92 pagesThe Pale Blood HuntRafael JunioNo ratings yet
- Introduction To Different Resources of Bioinformatics and Application PDFDocument55 pagesIntroduction To Different Resources of Bioinformatics and Application PDFSir RutherfordNo ratings yet
- Standards For Accreditation-Schools - 2010Document43 pagesStandards For Accreditation-Schools - 2010Nayda Jiménez Pérez100% (1)
- PSC Question BankDocument9 pagesPSC Question BankFaisal qblp100% (29)
- Kardiomed-700-User ManualDocument87 pagesKardiomed-700-User ManualJulia TimakovaNo ratings yet
- Lecture Notes:: Accident and Incident InvestigationDocument10 pagesLecture Notes:: Accident and Incident InvestigationajayikayodeNo ratings yet
- Project Proposal: A. Project Tittle: "Operation Cleanliness"Document8 pagesProject Proposal: A. Project Tittle: "Operation Cleanliness"Mike Avila100% (1)
- 1PE 4 Q3 Answer Sheet March 2024Document3 pages1PE 4 Q3 Answer Sheet March 2024Chid CabanesasNo ratings yet
- 3f PDFDocument4 pages3f PDFYidne GeoNo ratings yet
- TPF00391-3 Hotline BrochureDocument6 pagesTPF00391-3 Hotline BrochureAphichetNo ratings yet
- Colgate Marketing ReportDocument16 pagesColgate Marketing ReportRaja GuruNo ratings yet
- 1053-Article Text-2175-1-10-20230227Document8 pages1053-Article Text-2175-1-10-20230227inNo ratings yet
- Matter For PGDIPC Course PromotionDocument1 pageMatter For PGDIPC Course PromotionAnkur VasudevaNo ratings yet
- Et CareDocument15 pagesEt CarePaulo GarciaNo ratings yet
- DSM 5Document33 pagesDSM 5Grace Marie100% (2)
- Sterile Potassium Chloride 10%, 15% and 20% W/V Concentrate For Solution For InfusionDocument1 pageSterile Potassium Chloride 10%, 15% and 20% W/V Concentrate For Solution For InfusionMatthew ParsonsNo ratings yet
- Wiley Veterinary GynaecologyDocument2 pagesWiley Veterinary Gynaecologysanath0% (1)
- Development and in Vitro-In Vivo Evaluation of Gastro Retentive Drug Delivery of Nizatidine Using Natural and Semi - Synthetic PolymersDocument17 pagesDevelopment and in Vitro-In Vivo Evaluation of Gastro Retentive Drug Delivery of Nizatidine Using Natural and Semi - Synthetic PolymersShyamlaNo ratings yet
- MLC Lean Bronze Prep ClassDocument2 pagesMLC Lean Bronze Prep ClassSalaNo ratings yet
- Republic of The Philippines Davao Oriental State College of Science and TechnologyDocument4 pagesRepublic of The Philippines Davao Oriental State College of Science and Technologydinheadnursing09No ratings yet