You might also like
- 9.12.15-Dr. Vinod Rangari BiodataDocument11 pages9.12.15-Dr. Vinod Rangari BiodataNovi LatifaNo ratings yet
- 2006-SKELTON Obesity and Its Therapy From Genes To Community ActionDocument17 pages2006-SKELTON Obesity and Its Therapy From Genes To Community ActionNovi LatifaNo ratings yet
- Lcms PapainDocument1 pageLcms PapainNovi LatifaNo ratings yet
- Mclaws 2015Document11 pagesMclaws 2015Novi LatifaNo ratings yet
- 2007-KUMAR Genomic Medicine A New Frontier of Medicine in The Twenty FirstDocument5 pages2007-KUMAR Genomic Medicine A New Frontier of Medicine in The Twenty FirstNovi LatifaNo ratings yet
- Toxins: Ricinus Communis Intoxications in Human and VeterinaryDocument41 pagesToxins: Ricinus Communis Intoxications in Human and VeterinaryNovi LatifaNo ratings yet
- Fakultas Kehutanan Univeritas Tanjungpura. Jalan Imam Bonjol Pontianak 78124 EmailDocument12 pagesFakultas Kehutanan Univeritas Tanjungpura. Jalan Imam Bonjol Pontianak 78124 EmailNovi LatifaNo ratings yet
- Analysis of B-Galactosidase Production and Their Genes of Two Strains of Lactobacillus BulgaricusDocument5 pagesAnalysis of B-Galactosidase Production and Their Genes of Two Strains of Lactobacillus BulgaricusNovi LatifaNo ratings yet
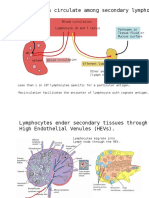
- Mature T Cells Circulate Among Secondary Lymphoid Tissues.: Blood CirculationDocument30 pagesMature T Cells Circulate Among Secondary Lymphoid Tissues.: Blood CirculationNovi LatifaNo ratings yet
- WellllDocument60 pagesWellllNovi LatifaNo ratings yet
- Cyanobacteria-Mediated Phenylpropanoids and Phytohormones in Rice (Oryza Sativa) Enhance Plant Growth and Stress ToleranceDocument12 pagesCyanobacteria-Mediated Phenylpropanoids and Phytohormones in Rice (Oryza Sativa) Enhance Plant Growth and Stress ToleranceNovi LatifaNo ratings yet
- Struktur & Fungsi Bunga, Buah, Dan Biji: Structure & Function Flower, Fruit, and SeedDocument13 pagesStruktur & Fungsi Bunga, Buah, Dan Biji: Structure & Function Flower, Fruit, and SeedNovi LatifaNo ratings yet
- Genome Constitution and Classification Using Retrotransposon-Based Markers in The Orphan Crop BananaDocument10 pagesGenome Constitution and Classification Using Retrotransposon-Based Markers in The Orphan Crop BananaNovi LatifaNo ratings yet
- Ravisha Raina Archit Sharma Saaksha Mantoo Arushee Mantoo Dr. Anita GrewalDocument3 pagesRavisha Raina Archit Sharma Saaksha Mantoo Arushee Mantoo Dr. Anita GrewalNovi LatifaNo ratings yet
- Art:10.1007/s00374 013 0840 4Document10 pagesArt:10.1007/s00374 013 0840 4Novi LatifaNo ratings yet
- Art:10 1007/BF01420232Document6 pagesArt:10 1007/BF01420232Novi LatifaNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Banana RND in MalaysiaDocument5 pagesBanana RND in MalaysiaChee'dida FelidaeNo ratings yet
- Farmer S Handbook On Banana Cultivars in The Philippines 1376Document78 pagesFarmer S Handbook On Banana Cultivars in The Philippines 1376atoydequitNo ratings yet
- Before Reading: A Threat To BananasDocument5 pagesBefore Reading: A Threat To BananasCCIEM CCIEMNo ratings yet
- Project Report On BananaDocument60 pagesProject Report On Bananaapi-26956361100% (1)
- In The Hands of Farm Workers: Can Banana Commercial Farms Survive? (2008)Document143 pagesIn The Hands of Farm Workers: Can Banana Commercial Farms Survive? (2008)laliga100% (1)
- UNIT 1 Test 2Document13 pagesUNIT 1 Test 2Howaida El MadawyNo ratings yet
- Banana - Wikipedia, The Free EncyclopediaDocument24 pagesBanana - Wikipedia, The Free EncyclopediaLeo ValentineNo ratings yet
- Project Report On BananaDocument64 pagesProject Report On Bananaapi-2695636194% (18)
- BananasDocument12 pagesBananasbookie183020% (1)
- Project Report On BananaDocument60 pagesProject Report On Bananavrushankhallayrock100% (1)
- E2 Mech English External Lab QPDocument6 pagesE2 Mech English External Lab QPGALI ANILKUMAR (N151229)No ratings yet