You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Physical Assessment Head To ToeDocument4 pagesPhysical Assessment Head To ToeFely Theresa Lanes Loreno95% (43)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Case Study 101: Abdominal Aortic Aneurysm With Acute Kidney InjuryDocument8 pagesCase Study 101: Abdominal Aortic Aneurysm With Acute Kidney InjuryPatricia Ann Nicole ReyesNo ratings yet
- Case 4 Reproductive Block PBLDocument9 pagesCase 4 Reproductive Block PBLFarisNo ratings yet
- GUIAS UCI para La Alimentación de Paciente CriticoDocument33 pagesGUIAS UCI para La Alimentación de Paciente CriticoFredy UmbarilaNo ratings yet
- S.N. System ID Name Speciality Guide: School of Nursing and ResearchDocument6 pagesS.N. System ID Name Speciality Guide: School of Nursing and ResearchBhawna JoshiNo ratings yet
- Practical Cardiology PDF 3vl DR NotesDocument709 pagesPractical Cardiology PDF 3vl DR NotesRemzi BoraNo ratings yet
- Practical Guide To Insulin Therapy in Type 2 Diabetes: January 2011Document91 pagesPractical Guide To Insulin Therapy in Type 2 Diabetes: January 2011leeyie22No ratings yet
- Norbert - Müller Inmunología y Psiquiatría PDFDocument411 pagesNorbert - Müller Inmunología y Psiquiatría PDFIvan Toscani100% (1)
- HypermagnesemiaDocument2 pagesHypermagnesemiaapi-3712326100% (1)
- Collarbone Breathing TreatmentDocument4 pagesCollarbone Breathing TreatmentPatrizioNo ratings yet
- 03 NCP 5 Pulmonary Tuberculosis LalitDocument17 pages03 NCP 5 Pulmonary Tuberculosis Lalitamit100% (2)
- SJ BDJ 2014 1053Document6 pagesSJ BDJ 2014 1053egray797No ratings yet
- Vestibular Function TestsDocument11 pagesVestibular Function TestsM. Arief Purnanta0% (1)
- Kenyaemr V17.3.1 Release Notes: Integration With Eid, Adt & Differential Upload DescriptionDocument3 pagesKenyaemr V17.3.1 Release Notes: Integration With Eid, Adt & Differential Upload DescriptionMigori Art DataNo ratings yet
- Acute Limb Ischemia Acute Limb Ischemia N SuddenDocument43 pagesAcute Limb Ischemia Acute Limb Ischemia N SuddenSWETHANo ratings yet
- Dead Space ManagementDocument26 pagesDead Space ManagementIgnasNo ratings yet
- Diagnosis and Management of Vertebral Compression FracturesDocument7 pagesDiagnosis and Management of Vertebral Compression FracturesMayang CendikiaNo ratings yet
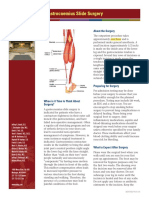
- Gastrocnemius Slide SurgeryDocument2 pagesGastrocnemius Slide SurgeryHein Aung ZawNo ratings yet
- Problem 1 Emergency Medicine Block: Agustina Cynthia Cesari S 405140066 Group 1Document90 pagesProblem 1 Emergency Medicine Block: Agustina Cynthia Cesari S 405140066 Group 1Astri GunardiNo ratings yet
- General AnesthesiaDocument22 pagesGeneral AnesthesiaDr Anais AsimNo ratings yet
- A Special Interview With Dr. Meryl Nass by Dr. Joseph MercolaDocument9 pagesA Special Interview With Dr. Meryl Nass by Dr. Joseph MercolaJonathan Robert Kraus (OutofMudProductions)No ratings yet
- OP 4118 Root Cause AnalysisDocument16 pagesOP 4118 Root Cause AnalysisAhmed Hanno100% (1)
- HistologyDocument15 pagesHistologySara ŠvedekNo ratings yet
- Reviewer RenalDocument13 pagesReviewer Renalsean blaze100% (1)
- Case Write-Up 2Document22 pagesCase Write-Up 2Syed TalhaNo ratings yet
- Etiology of Tuberculosis PDFDocument2 pagesEtiology of Tuberculosis PDFKevinNo ratings yet
- Fracture Healing - Basic Science - Orthobullets PDFDocument2 pagesFracture Healing - Basic Science - Orthobullets PDFMelAcostaNo ratings yet
- Janet RowleyDocument1 pageJanet RowleyANA GABRIELA PEREZ VIDALNo ratings yet
- HydrotherapyDocument40 pagesHydrotherapyThopu UmamaheswariNo ratings yet
- Barodontalgia As A Differential Diagnosis PDFDocument4 pagesBarodontalgia As A Differential Diagnosis PDFDavid ColonNo ratings yet