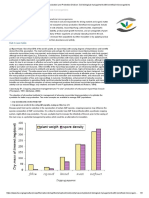
You might also like
- Microbial FertilizersDocument7 pagesMicrobial FertilizersSakthi PNo ratings yet
- Pub Microbial Strategies For Crop ImprovementDocument371 pagesPub Microbial Strategies For Crop Improvementamrita kanuNo ratings yet
- Introduction:-: Bio Fertilizers: An Alternative of Chemical FertilizersDocument26 pagesIntroduction:-: Bio Fertilizers: An Alternative of Chemical FertilizersRachna chaturvediNo ratings yet
- Article 1 Biofertilizers and Sustainable AgricultureDocument12 pagesArticle 1 Biofertilizers and Sustainable AgriculturePaolita CortesNo ratings yet
- Microbial Ecology and Diversity AssignmentDocument19 pagesMicrobial Ecology and Diversity AssignmentGutu EmiruNo ratings yet
- tmpD493 TMPDocument11 pagestmpD493 TMPFrontiersNo ratings yet
- Sustainability 13 01868 v2Document20 pagesSustainability 13 01868 v2Maria Angelica Barrera OspinaNo ratings yet
- Plant Growth Promoting Bacteria As An Alternative Strategy For Salt ToleranceDocument12 pagesPlant Growth Promoting Bacteria As An Alternative Strategy For Salt ToleranceRiz Jomaica ArenasNo ratings yet
- M C Jaizme Vega Et Al - Effect of Arbuscular Mycorrhizal Fungi (Amf) and Other Rhizosphere Microorganisms On Development of The Banana Root SystemDocument15 pagesM C Jaizme Vega Et Al - Effect of Arbuscular Mycorrhizal Fungi (Amf) and Other Rhizosphere Microorganisms On Development of The Banana Root SystemandresuribeNo ratings yet
- Aarti YadavDocument5 pagesAarti YadavIshitaNo ratings yet
- Kumar 2016Document16 pagesKumar 2016Gustavo Cano GallegosNo ratings yet
- Biocatalysis and Agricultural BiotechnologyDocument12 pagesBiocatalysis and Agricultural BiotechnologyMufidatun Nafi'ahNo ratings yet
- 1 ACapstoneeeeersDocument30 pages1 ACapstoneeeeersSamantha Jeka Bianca Asna100% (2)
- Babalola 2010 Beneficial Bacteria of Agricultural ImportanceDocument12 pagesBabalola 2010 Beneficial Bacteria of Agricultural ImportanceCarlos AndradeNo ratings yet
- Bacterial Siderophoreasa Plant Growth PROMOTERDocument18 pagesBacterial Siderophoreasa Plant Growth PROMOTEREdgar Meneses MoranNo ratings yet
- Plant Growth Promoting RhizobiaDocument23 pagesPlant Growth Promoting RhizobiadanyjorgeNo ratings yet
- Beneficial Bacteria of Agricultural ImportanceDocument12 pagesBeneficial Bacteria of Agricultural ImportanceXevin MoreiraNo ratings yet
- Mechanisms and Applications of Plant Growth Promoting RhizobacteriaDocument48 pagesMechanisms and Applications of Plant Growth Promoting RhizobacteriaFabricio Carriel CoelloNo ratings yet
- Ac 122 Sir PortionDocument64 pagesAc 122 Sir PortionjyotiranjanNo ratings yet
- Plant Production and Protection Division - Soil Biological Management With Beneficial MicroorganismsDocument3 pagesPlant Production and Protection Division - Soil Biological Management With Beneficial MicroorganismsCyberSquare IndiaNo ratings yet
- Bio FertilizerDocument35 pagesBio FertilizerkrithideepNo ratings yet
- The Comparative Study of Biofertilizers On Growth and Yield of Wheat: A ReviewDocument6 pagesThe Comparative Study of Biofertilizers On Growth and Yield of Wheat: A ReviewInternational Journal of Innovative Science and Research Technology100% (1)
- Progress in Microbial Fertilizer Regulation of Crop Growth and Soil-2024Document21 pagesProgress in Microbial Fertilizer Regulation of Crop Growth and Soil-2024Adios ANo ratings yet
- 1 s2.0 S240584402203897X MainDocument13 pages1 s2.0 S240584402203897X MainEstefania GarciaNo ratings yet
- Biocatalysis and Agricultural BiotechnologyDocument11 pagesBiocatalysis and Agricultural BiotechnologyDr Arunprasath ANo ratings yet
- DiversityDocument25 pagesDiversityK. Jeremy Hidalgo ArnezNo ratings yet
- Unlocking The Potential of Plant Growth-Promoting Rhizobacteria On Soil Health and The Sustainability of Agricultural SystemsDocument20 pagesUnlocking The Potential of Plant Growth-Promoting Rhizobacteria On Soil Health and The Sustainability of Agricultural Systemsmazhar.commereceNo ratings yet
- Chapter 2-REVIEW OF RELATED LITERATUREDocument18 pagesChapter 2-REVIEW OF RELATED LITERATUREMaria Lovella Misolas Diaz100% (1)
- Colecciones Microbianas y Su Potencial en La Seguridad AlimentariaDocument8 pagesColecciones Microbianas y Su Potencial en La Seguridad AlimentariaSecuenciación NomadNo ratings yet
- Nitrogen Is One of The Essential Elements Required For The Synthesis of Amino Acids WhichDocument11 pagesNitrogen Is One of The Essential Elements Required For The Synthesis of Amino Acids Whichsanjay kumarNo ratings yet
- PGPRs ReviewDocument15 pagesPGPRs ReviewMuhammad Asif RoyNo ratings yet
- Acta Micro - 2022 - Amrita PGPR BiofertilizerDocument10 pagesActa Micro - 2022 - Amrita PGPR BiofertilizerSarita Sanjay SharmaNo ratings yet
- Bio-Fertilizer A Highly Potent AlternatiDocument14 pagesBio-Fertilizer A Highly Potent AlternatiSandra Magally Sanchez TrujilloNo ratings yet
- Agronomy 10 01415Document12 pagesAgronomy 10 01415AB LimaNo ratings yet
- Crop Soil OrganismsDocument16 pagesCrop Soil OrganismsJohn Eric CabañaNo ratings yet
- Bio BoosterDocument3 pagesBio BoosterUmarqayum Abu BakarNo ratings yet
- Soil Microbial Resources For Improving Fertilizers Efficiency in An Integrated Plant Nutrient Management SystemDocument25 pagesSoil Microbial Resources For Improving Fertilizers Efficiency in An Integrated Plant Nutrient Management SystemRifqi Fathul ArroisiNo ratings yet
- Biofertilizer - WikipediaDocument24 pagesBiofertilizer - Wikipedia30521u33009No ratings yet
- BiofertilizersDocument8 pagesBiofertilizersUmer FarooqNo ratings yet
- Screening of Rhizospheric Bacteria That Plays Role in Increasing Plant GrowthDocument15 pagesScreening of Rhizospheric Bacteria That Plays Role in Increasing Plant Growthpratiwi kusumaNo ratings yet
- TERRESTIAL Eco proposalWITHDATAFORANALYSISDocument22 pagesTERRESTIAL Eco proposalWITHDATAFORANALYSISShiela PlanasNo ratings yet
- NTCC DeepakDocument21 pagesNTCC DeepakDeepak KumarNo ratings yet
- Agriculture 09 00122 v3Document13 pagesAgriculture 09 00122 v3dinchus_106No ratings yet
- Communication in The Rhizosphere, A Target For Pest ManagementDocument25 pagesCommunication in The Rhizosphere, A Target For Pest ManagementHector Chavez TeuberNo ratings yet
- The Occurrence of Pesticides in Environment and Current Technologies For Their Remediation and ManagementDocument36 pagesThe Occurrence of Pesticides in Environment and Current Technologies For Their Remediation and ManagementIJAR JOURNALNo ratings yet
- Agrultura MicrobiosDocument25 pagesAgrultura Microbioschobyts27lizyNo ratings yet
- Chapter I IntroductionDocument6 pagesChapter I IntroductionShubham BorkarNo ratings yet
- A Review Biofertilizer A Key Player in Enhancing Soil Fertility and CropproductivityDocument7 pagesA Review Biofertilizer A Key Player in Enhancing Soil Fertility and CropproductivitySUNILNo ratings yet
- Trichoderma Species: Versatile Plant SymbiontsDocument11 pagesTrichoderma Species: Versatile Plant SymbiontsAdam StankiewiczNo ratings yet
- The Role of Organic Acids On Microbial Deterioration in The Radix Continuous Monoculture RegimesDocument13 pagesThe Role of Organic Acids On Microbial Deterioration in The Radix Continuous Monoculture RegimesWilly GonzalesNo ratings yet
- tmpAEC8 TMPDocument12 pagestmpAEC8 TMPFrontiersNo ratings yet
- J. Inter Mikroba Tanah 2Document10 pagesJ. Inter Mikroba Tanah 2Fawkes LightNo ratings yet
- Bacteria Fungi and Archaea Domains in Rhizospheric Soil and Their Effects in Enhancing Agricultural ProductivityDocument20 pagesBacteria Fungi and Archaea Domains in Rhizospheric Soil and Their Effects in Enhancing Agricultural ProductivityFungicolas JaverianaNo ratings yet
- Artículo AIMS Microbiology 3. 2017Document23 pagesArtículo AIMS Microbiology 3. 2017Aquiles18No ratings yet
- 10 1002@jobm 202000370Document34 pages10 1002@jobm 202000370Tahiri AbdelilahNo ratings yet
- Amino Chelate Fertilizers As The Latest Novelties in Plant Nutrition A ReviewDocument4 pagesAmino Chelate Fertilizers As The Latest Novelties in Plant Nutrition A ReviewEditor IJTSRDNo ratings yet
- GJESM Volume 7 Issue 3 Pages 401-418Document18 pagesGJESM Volume 7 Issue 3 Pages 401-418GJESMNo ratings yet
- High-Throughput Customization of Plant Microbiomes For Sustainable AgricultureDocument9 pagesHigh-Throughput Customization of Plant Microbiomes For Sustainable AgricultureNeta Adzkiya HamidahNo ratings yet
- Litreture ReviewDocument4 pagesLitreture ReviewWalter HudsonNo ratings yet
- Functional Diversity of Mycorrhiza and Sustainable Agriculture: Management to Overcome Biotic and Abiotic StressesFrom EverandFunctional Diversity of Mycorrhiza and Sustainable Agriculture: Management to Overcome Biotic and Abiotic StressesNo ratings yet
- ConstructionDocument13 pagesConstructionJoão RodriguesNo ratings yet
- Chapter 05Document10 pagesChapter 05João RodriguesNo ratings yet
- Chapter 10Document9 pagesChapter 10João RodriguesNo ratings yet
- Chapter 09Document9 pagesChapter 09João RodriguesNo ratings yet
- Chapter 06Document3 pagesChapter 06João RodriguesNo ratings yet
- Chapter 04Document8 pagesChapter 04João RodriguesNo ratings yet
- Chapter 08Document12 pagesChapter 08João RodriguesNo ratings yet
- Chapter 07Document11 pagesChapter 07jacquesjzaNo ratings yet
- Appendix 6Document5 pagesAppendix 6João RodriguesNo ratings yet
- Chapter 01Document16 pagesChapter 01freakyarch4290No ratings yet
- Passive DesignDocument21 pagesPassive DesignpassionpropelNo ratings yet
- Passive DesignDocument11 pagesPassive DesignpassionpropelNo ratings yet
- Appendix 4 U-Values For Windows With Insulating CoversDocument1 pageAppendix 4 U-Values For Windows With Insulating CoversJoão RodriguesNo ratings yet
- Appendix 5Document6 pagesAppendix 5João RodriguesNo ratings yet
- Hacker Inside Vol 1Document97 pagesHacker Inside Vol 1Claudemir Resende100% (1)
- Hacker Inside Vol.2Document88 pagesHacker Inside Vol.2Miguel Machado GonçalvesNo ratings yet
- Appendix 1Document3 pagesAppendix 1Piyush SardaNo ratings yet
- Appendix 3 Maps of The Average Percentage of The Time The Sun Is ShiningDocument5 pagesAppendix 3 Maps of The Average Percentage of The Time The Sun Is ShiningJoão RodriguesNo ratings yet
- Appendix 2 Solar Radiation On South Walls On Sunny DaysDocument4 pagesAppendix 2 Solar Radiation On South Walls On Sunny DaysJoão RodriguesNo ratings yet
- Appendix 2 Solar Radiation On South Walls On Sunny DaysDocument4 pagesAppendix 2 Solar Radiation On South Walls On Sunny DaysJoão RodriguesNo ratings yet
- US Army Medical Course MD0180-100 - Radio Logical HealthDocument80 pagesUS Army Medical Course MD0180-100 - Radio Logical HealthGeorges100% (2)
- Hacker Inside Vol.3Document69 pagesHacker Inside Vol.3Miguel Machado GonçalvesNo ratings yet
- Hacker Inside Vol.4 WWW - PiratatugaDocument88 pagesHacker Inside Vol.4 WWW - PiratatugaNick SlimNo ratings yet
- Atlas of Caesium Deposition On Europe After The Chernobyl Accident by European Communities 928283140xDocument65 pagesAtlas of Caesium Deposition On Europe After The Chernobyl Accident by European Communities 928283140xJoão Rodrigues100% (1)
- Acupuncture For PhysiotherapistDocument277 pagesAcupuncture For PhysiotherapistLuis Gomes100% (1)
- Hacker Inside - Vol. 5Document75 pagesHacker Inside - Vol. 5Daniel MetamorphNo ratings yet
- Acupuncture, Trigger Points & Musculoskeletal PainDocument362 pagesAcupuncture, Trigger Points & Musculoskeletal PainThezza Lion100% (2)
- Essentials of Chinese AcupunctureDocument448 pagesEssentials of Chinese AcupunctureShea-Baby Sheabutterproducts100% (2)
- Acupuncture in Practice by Anthony CampbellDocument179 pagesAcupuncture in Practice by Anthony CampbellRuth Manel100% (13)
- Veterinary AcupunctureDocument305 pagesVeterinary AcupunctureMihalache Alex100% (7)
- Research Article Arbuscular Mycorrhiza Improves Yield and Nutritional Properties of Onion (Allium Cepa)Document21 pagesResearch Article Arbuscular Mycorrhiza Improves Yield and Nutritional Properties of Onion (Allium Cepa)UmiChabibahNo ratings yet
- AMF and Soil FertilityDocument15 pagesAMF and Soil FertilityCharu KaushikNo ratings yet
- Fusconi (1983) Reported That The Short Root System PlantsDocument16 pagesFusconi (1983) Reported That The Short Root System Plantsكيرلس مجدىNo ratings yet
- FINAL-PAPER (Eto Na Talaga)Document74 pagesFINAL-PAPER (Eto Na Talaga)Raymund Arcos100% (1)
- Taming The Truffle References Edited Plus Additions PDFDocument42 pagesTaming The Truffle References Edited Plus Additions PDFBogdana BogdanNo ratings yet
- Dictionary of Ecology and EnvironmentalDocument170 pagesDictionary of Ecology and EnvironmentalChristian TambacNo ratings yet
- Biodiversity and Ecosystem ServicesDocument18 pagesBiodiversity and Ecosystem Servicessameer AhadNo ratings yet
- Jayraj Thesis Okra PDFDocument127 pagesJayraj Thesis Okra PDFMohitNo ratings yet
- Handbook of Vegetation ScienceDocument378 pagesHandbook of Vegetation ScienceSergio Valdivia RiveraNo ratings yet
- AmfDocument165 pagesAmfJesus OcampoNo ratings yet
- VAMDocument1 pageVAMcnu0294No ratings yet
- 81 2446 4 PBDocument9 pages81 2446 4 PBpetrus alkantaraNo ratings yet
- Review of LiteratureDocument15 pagesReview of LiteratureMuhammad Ali Khan75% (8)
- Value-Added Products of Rice Husk in Various DisciplinesDocument17 pagesValue-Added Products of Rice Husk in Various DisciplinesIJRASETPublicationsNo ratings yet
- Mycorrhiza AssignmentDocument4 pagesMycorrhiza AssignmentmohkristNo ratings yet
- Kumar 2016Document16 pagesKumar 2016Gustavo Cano GallegosNo ratings yet
- Factors Affecting Crop ProductionDocument36 pagesFactors Affecting Crop ProductionPca ZdnNo ratings yet
- SoilDocument9 pagesSoilsoumyarm942No ratings yet
- Mass Production of AM Fungal Inoculum by Soil Based Pot CultureDocument5 pagesMass Production of AM Fungal Inoculum by Soil Based Pot CultureIJARBS JOURNALNo ratings yet
- 01 SidhoumDocument12 pages01 SidhoumFilip MartinovićNo ratings yet
- Mycorrhiza 180310082441Document58 pagesMycorrhiza 180310082441Khadija100% (1)
- Seleksi Lima Isolat Fungi Mikoriza Arbuskular Untuk Pembibitan Kelapa Sawit (Elaeis Guineensis Jacq.) Pada Dua Dosis Pupuk NPKDocument9 pagesSeleksi Lima Isolat Fungi Mikoriza Arbuskular Untuk Pembibitan Kelapa Sawit (Elaeis Guineensis Jacq.) Pada Dua Dosis Pupuk NPKsitajamNo ratings yet
- Soil Fertility and Plant NutritionpptDocument221 pagesSoil Fertility and Plant NutritionpptTäð Œvê MîðNo ratings yet
- Plant Archives: Number 2 October - 2019Document18 pagesPlant Archives: Number 2 October - 2019SAADNo ratings yet
- Biofertilizers TnauDocument17 pagesBiofertilizers TnaudonbosskissNo ratings yet
- Types of Biofertilizers and Importance: I. N FixersDocument12 pagesTypes of Biofertilizers and Importance: I. N FixersshubhamNo ratings yet
- Maximum Yield USA June 2017 PDFDocument166 pagesMaximum Yield USA June 2017 PDFmushroomman88No ratings yet
- Efektifitas Fungi Mikoriza Arbuskula (Fma) Serta Pupuk Nitrogen Dan Fosfat Terhadap PERTUMBUHAN SEMAI JABON (Anthocephallus Cadamba Roxb.)Document6 pagesEfektifitas Fungi Mikoriza Arbuskula (Fma) Serta Pupuk Nitrogen Dan Fosfat Terhadap PERTUMBUHAN SEMAI JABON (Anthocephallus Cadamba Roxb.)Raudhatul RahmahNo ratings yet
- Arbuscular Mycorrhizae As A Component of INM - 2Document11 pagesArbuscular Mycorrhizae As A Component of INM - 2Rizky SanjayaNo ratings yet
- Sundarban Mangrove Plants & Am FungiDocument13 pagesSundarban Mangrove Plants & Am FungiAvigyan ChatterjeeNo ratings yet