You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Volvo A30e2 PDFDocument28 pagesVolvo A30e2 PDFsafdey100% (1)
- (QUIZ) NEMATODES (2nd Year BS Medical Technology)Document2 pages(QUIZ) NEMATODES (2nd Year BS Medical Technology)avery aerNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Ecs Omsm (Operation, Maintenance & Safety Manual) : Electro-Cleen™ SystemDocument233 pagesEcs Omsm (Operation, Maintenance & Safety Manual) : Electro-Cleen™ SystemElena Gaevska100% (1)
- Review of Related Literature of Jackfruit SeedDocument4 pagesReview of Related Literature of Jackfruit SeedPrince Martin John Villamor80% (5)
- TRALEG KYABGON - The-Influence-of-Yogacara-on-Mahamudra PDFDocument145 pagesTRALEG KYABGON - The-Influence-of-Yogacara-on-Mahamudra PDFFilippo Lunardo100% (1)
- Kohler RRT ManualDocument40 pagesKohler RRT Manualjmh488100% (1)
- Reading and Interpreting Product DesignDocument38 pagesReading and Interpreting Product DesignKate Adriano100% (2)
- First Periodical Mapeh 9Document5 pagesFirst Periodical Mapeh 9Juvy Singian100% (1)
- NSL Hollow Core Slabs 12jul16 - 2016 Edition PDFDocument24 pagesNSL Hollow Core Slabs 12jul16 - 2016 Edition PDFjohnkoh9950% (2)
- COMMUNITY HEALTH NURSING: PREVENTION, FAMILIES, PRIMARY CAREDocument4 pagesCOMMUNITY HEALTH NURSING: PREVENTION, FAMILIES, PRIMARY CAREJohn Vincent VasquezNo ratings yet
- Marine Debris and Human Impacts On Sea Turtles in Southern BrazilDocument5 pagesMarine Debris and Human Impacts On Sea Turtles in Southern BrazilIlham ClinkersNo ratings yet
- 09 Suhartono NurdinDocument8 pages09 Suhartono NurdinIlham ClinkersNo ratings yet
- Nilai Planton 2017Document5 pagesNilai Planton 2017Ilham ClinkersNo ratings yet
- 124-From Mangroves To Coral Reefs Partie2Document57 pages124-From Mangroves To Coral Reefs Partie2Ilham ClinkersNo ratings yet
- Jurnal Pengaruh Derajat Keasaman (PH) Air Laut Terhadap Konsentrasi Kalsium Dan Laju Pertumbuhan HalimedaDocument8 pagesJurnal Pengaruh Derajat Keasaman (PH) Air Laut Terhadap Konsentrasi Kalsium Dan Laju Pertumbuhan HalimedaRobiansyahNo ratings yet
- Jurnal Pengaruh Derajat Keasaman (PH) Air Laut Terhadap Konsentrasi Kalsium Dan Laju Pertumbuhan HalimedaDocument8 pagesJurnal Pengaruh Derajat Keasaman (PH) Air Laut Terhadap Konsentrasi Kalsium Dan Laju Pertumbuhan HalimedaRobiansyahNo ratings yet
- 124-From Mangroves To Coral Reefs Partie2Document57 pages124-From Mangroves To Coral Reefs Partie2Ilham ClinkersNo ratings yet
- Review Paper 1Document10 pagesReview Paper 1Ilham ClinkersNo ratings yet
- 8 1 1958 FishingDocument8 pages8 1 1958 FishingIlham ClinkersNo ratings yet
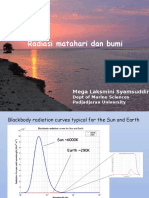
- Radiasi Matahari Dan BumiDocument14 pagesRadiasi Matahari Dan BumiIlham ClinkersNo ratings yet
- 1 s2.0 S0006320714004212 MainDocument13 pages1 s2.0 S0006320714004212 MainIlham ClinkersNo ratings yet
- Deteksi BiosonarDocument7 pagesDeteksi BiosonarIlham ClinkersNo ratings yet
- Mammals at Large Spatial ScalesDocument17 pagesMammals at Large Spatial ScalesIlham ClinkersNo ratings yet
- Colour Vision in Aquatic Mammals-Facts and Open Questions: Ulrike Griebel and Leo PeichlDocument13 pagesColour Vision in Aquatic Mammals-Facts and Open Questions: Ulrike Griebel and Leo PeichlIlham ClinkersNo ratings yet
- Deskripsi Anatomi of DolphinDocument12 pagesDeskripsi Anatomi of DolphinIlham ClinkersNo ratings yet
- Local Wisdom-Based Curriculum for Standardization EducationDocument17 pagesLocal Wisdom-Based Curriculum for Standardization EducationIlham ClinkersNo ratings yet
- Motion Detection Application Using Web CameraDocument3 pagesMotion Detection Application Using Web CameraIlham ClinkersNo ratings yet
- Colour Vision in Aquatic Mammals-Facts and Open Questions: Ulrike Griebel and Leo PeichlDocument13 pagesColour Vision in Aquatic Mammals-Facts and Open Questions: Ulrike Griebel and Leo PeichlIlham ClinkersNo ratings yet
- Deskripsi Anatomi of DolphinDocument12 pagesDeskripsi Anatomi of DolphinIlham ClinkersNo ratings yet
- Food Sharing DolphinsDocument5 pagesFood Sharing DolphinsIlham ClinkersNo ratings yet
- Agresi Jantan Dolphins PDFDocument3 pagesAgresi Jantan Dolphins PDFIlham ClinkersNo ratings yet
- Alloparental DolphinEDocument5 pagesAlloparental DolphinEIlham ClinkersNo ratings yet
- Belecobeauty Company ProfileDocument19 pagesBelecobeauty Company ProfileBisma BrawijayaNo ratings yet
- Cardiovascular SystemDocument40 pagesCardiovascular SystemDouglas Jacques100% (1)
- The Journey of HIV 1 Non-Nucleoside Reverse TranscriptaseDocument33 pagesThe Journey of HIV 1 Non-Nucleoside Reverse TranscriptaseAnkit Kumar SinghNo ratings yet
- DIFFERENTIALDocument4 pagesDIFFERENTIALsaffrinNo ratings yet
- Computer PackagesDocument72 pagesComputer PackagesBildad JoashNo ratings yet
- Noon Fees StructureDocument8 pagesNoon Fees StructureNithin SreekumarNo ratings yet
- ASTM A193-A193M-04cDocument12 pagesASTM A193-A193M-04cNadhiraNo ratings yet
- Heat Transfer Through Extended SurfacesDocument16 pagesHeat Transfer Through Extended SurfaceschawarepNo ratings yet
- SEISMIC LOAD ANALYSISDocument9 pagesSEISMIC LOAD ANALYSISAditya RoyNo ratings yet
- Incepbio Project PPT - 080124Document21 pagesIncepbio Project PPT - 080124adminpuneNo ratings yet
- DMAE Powder Safety Data SheetDocument3 pagesDMAE Powder Safety Data SheetAInhoaNo ratings yet
- Deka Batteries PDFDocument6 pagesDeka Batteries PDFLuis Alonso Osorio MolinaNo ratings yet
- 10TH Gseb PaperDocument10 pages10TH Gseb Paperkhushboo guptaNo ratings yet
- Clerical Exam Sample PaperDocument21 pagesClerical Exam Sample PaperSarbjit Singh100% (1)
- International TradeDocument28 pagesInternational TradeAmit Sharma0% (1)
- Assignment On Computer HardwareDocument9 pagesAssignment On Computer HardwareMuktadirhasan100% (1)
- Unit 4 Out WorldDocument45 pagesUnit 4 Out WorldKhánh Nguyễn100% (2)
- Balzac GaitDocument7 pagesBalzac Gaithieratic_headNo ratings yet
- q4_tleDocument65 pagesq4_tleAngelica TaerNo ratings yet
- Report on internship at JDS TransformersDocument50 pagesReport on internship at JDS TransformersSachinNo ratings yet