You might also like
- The Structure of A Kidney NephronDocument4 pagesThe Structure of A Kidney NephronMarie St. Louis100% (1)
- Org Paper Draft-1scribdDocument2 pagesOrg Paper Draft-1scribdMarie St. LouisNo ratings yet
- Vision LectureDocument24 pagesVision LectureMarie St. Louis100% (1)
- US Internal Revenue Service: f2106 AccessibleDocument2 pagesUS Internal Revenue Service: f2106 AccessibleIRS100% (1)
- Vision Lecture IIDocument12 pagesVision Lecture IIMarie St. LouisNo ratings yet
- Org Paper Draft-1scribdDocument2 pagesOrg Paper Draft-1scribdMarie St. LouisNo ratings yet
- EpitheliumDocument10 pagesEpitheliumMarie St. Louis100% (1)
- Vision and MemoryDocument10 pagesVision and MemoryMarie St. LouisNo ratings yet
- Eye Memory VisonDocument15 pagesEye Memory VisonMarie St. LouisNo ratings yet
- EyeDocument3 pagesEyeMarie St. LouisNo ratings yet
- Vision Powerpoint IDocument22 pagesVision Powerpoint IMarie St. Louis100% (1)
- Eye and FeatureDocument16 pagesEye and FeatureMarie St. LouisNo ratings yet
- HowDocument8 pagesHowMarie St. LouisNo ratings yet
- msl302 L11a Motivating Soldiers SlidesDocument16 pagesmsl302 L11a Motivating Soldiers SlidesMarie St. LouisNo ratings yet
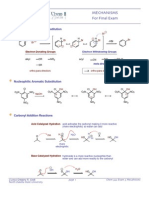
- Mechanisms For Final Exam: Electrophilic Aromatic SubstitutionDocument4 pagesMechanisms For Final Exam: Electrophilic Aromatic SubstitutionMarie St. LouisNo ratings yet
- msl302 l11c Consideration of Others SlidesDocument44 pagesmsl302 l11c Consideration of Others SlidesMarie St. LouisNo ratings yet
- NMR 13c 05Document34 pagesNMR 13c 05Marie St. LouisNo ratings yet
- Organic Chem Reactions Mindmap PDFDocument7 pagesOrganic Chem Reactions Mindmap PDFgsbabil100% (1)
- Sexual HarrassementDocument10 pagesSexual HarrassementMarie St. LouisNo ratings yet
- Summary of Organic ReactionsDocument21 pagesSummary of Organic ReactionsMarie St. LouisNo ratings yet
- Finalreview 03Document30 pagesFinalreview 03Marie St. LouisNo ratings yet
- Organic Chemistry 342 ReactionsDocument11 pagesOrganic Chemistry 342 ReactionsFrancisco QuirozNo ratings yet
- Organic Chemistry Final Exam KeyDocument10 pagesOrganic Chemistry Final Exam KeybrookNo ratings yet
- Letter To Creditor For Reduction of DebtDocument4 pagesLetter To Creditor For Reduction of DebtMarie St. Louis0% (1)
- Cells of Erogenous Zones Powerpoint FinalDocument32 pagesCells of Erogenous Zones Powerpoint FinalMarie St. Louis100% (1)
- Restriction Enzyme LabDocument12 pagesRestriction Enzyme LabMarie St. LouisNo ratings yet
- Cell DivisionDocument8 pagesCell DivisionMarie St. Louis100% (1)
- Debora Biomass and Alternative FuelDocument58 pagesDebora Biomass and Alternative FuelMarie St. LouisNo ratings yet
- Water PowerpointDocument16 pagesWater PowerpointMarie St. LouisNo ratings yet
- Res Enzyme Lab IIDocument2 pagesRes Enzyme Lab IIMarie St. LouisNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Central Dogma of Molecular BiologyDocument54 pagesCentral Dogma of Molecular BiologyQuintero MerylNo ratings yet
- Ligation Sequencing gDNA V14 - Whole Genome Amplification (SQK-LSK114) - MinionDocument8 pagesLigation Sequencing gDNA V14 - Whole Genome Amplification (SQK-LSK114) - MinionDevansh PanwarNo ratings yet
- Avoiding Depurination During Trityl-On Purification: Author: Greg ScottDocument8 pagesAvoiding Depurination During Trityl-On Purification: Author: Greg ScottKasi RuddrarajuNo ratings yet
- Protein Synthesis Worksheet PART A. Read The Following and Take Notes On Your PaperDocument2 pagesProtein Synthesis Worksheet PART A. Read The Following and Take Notes On Your PaperLemonade BlinkNo ratings yet
- A Basic PCR ProtocolDocument4 pagesA Basic PCR ProtocolIrfan HussainNo ratings yet
- PCR GelDocDocument15 pagesPCR GelDocKean GBNo ratings yet
- DNA Modifying EnzymesDocument26 pagesDNA Modifying EnzymesShikha YashveerNo ratings yet
- Dna, Rna, Transcription, Translation & Protein Synthesis PDFDocument14 pagesDna, Rna, Transcription, Translation & Protein Synthesis PDFSaeed Khan100% (1)
- Understanding Your Mtdna Results: My Ftdna CodeDocument4 pagesUnderstanding Your Mtdna Results: My Ftdna Code1sniper1No ratings yet
- DNA Replication Escape Room DocumentsDocument42 pagesDNA Replication Escape Room DocumentsVictoria WarrenNo ratings yet
- Structure of DNA Exam Questions and Mark Scheme 24lwmg4Document12 pagesStructure of DNA Exam Questions and Mark Scheme 24lwmg4andre gironNo ratings yet
- Long Test As Level Nucleic AcidDocument9 pagesLong Test As Level Nucleic AcidPrisky Jofan SimbarNo ratings yet
- 2013-MiRNA Regulatory Variation in Human EvolutionDocument9 pages2013-MiRNA Regulatory Variation in Human EvolutionJorge Hantar Touma LazoNo ratings yet
- Small Molecule Tools To ModulaDocument329 pagesSmall Molecule Tools To Modula吴善统No ratings yet
- Bio 2Document3 pagesBio 2ganchimeg gankhuuNo ratings yet
- Gene SilencingDocument18 pagesGene SilencingNAMRA RASHEEDNo ratings yet
- DNA Structure and Organization in The CellDocument39 pagesDNA Structure and Organization in The CellRyan Wilson - Sandalwood Heights SS (2442)No ratings yet
- Truseq Stranded Totalrna Sampleprep GuideDocument162 pagesTruseq Stranded Totalrna Sampleprep GuideVinicius Ferreira da PaixãoNo ratings yet
- NEB IlluminaDocument13 pagesNEB IlluminaLei Zi DuNo ratings yet
- Preston1991Document5 pagesPreston1991Alberto TerronesNo ratings yet
- Telomere PDFDocument2 pagesTelomere PDFsoumita100% (1)
- Central Dogma of Life: Transcription and TranslationDocument13 pagesCentral Dogma of Life: Transcription and TranslationLamia AkterNo ratings yet
- Henry See 2102515J A21M4 MDT Lab ReportDocument12 pagesHenry See 2102515J A21M4 MDT Lab ReportHenry SeeNo ratings yet
- DV200 Technote Truseq Rna AccessDocument4 pagesDV200 Technote Truseq Rna AccessIoannis PanagopoulosNo ratings yet
- TranslationDocument51 pagesTranslationAleena MustafaNo ratings yet
- Oligonucleotide Synthesis Order FormDocument4 pagesOligonucleotide Synthesis Order FormUjwal TrivediNo ratings yet
- Cis I Trans ElementiDocument37 pagesCis I Trans ElementiHexaGlobinNo ratings yet
- Central Dogma of Molecular Biology RendraDocument36 pagesCentral Dogma of Molecular Biology RendraYuliaji Narendra PutraNo ratings yet
- Cytogenetics Topic 6 Hand OutDocument18 pagesCytogenetics Topic 6 Hand OutJean Carmelette BalalloNo ratings yet
- OC4 2-13 Automation GeethaDocument3 pagesOC4 2-13 Automation GeethaCarlos RamírezNo ratings yet