You might also like
- 1999, JCP REVIEW The Molecular Basis of Disorders of Red Cell EnzymesDocument4 pages1999, JCP REVIEW The Molecular Basis of Disorders of Red Cell EnzymesSukma EffendyNo ratings yet
- A6 MMWR Update On IGRA in Diagnosis of TB 2010Document28 pagesA6 MMWR Update On IGRA in Diagnosis of TB 2010Rhea DerijeNo ratings yet
- Cpns Soal2Document25 pagesCpns Soal2Sukma EffendyNo ratings yet
- 2010 Systematic Review Interferon Gamma Release Assays For Tuberculosis Screening of Healthcare WorkersDocument9 pages2010 Systematic Review Interferon Gamma Release Assays For Tuberculosis Screening of Healthcare WorkersSukma EffendyNo ratings yet
- Tanpa JudulDocument689 pagesTanpa JudulSukma Effendy100% (3)
- 2010 Research Quantitative Analysis of Urine Based Assay For Detection of LAM in Patient With TB PDFDocument3 pages2010 Research Quantitative Analysis of Urine Based Assay For Detection of LAM in Patient With TB PDFSukma EffendyNo ratings yet
- 032-Roubort-Advantages and Integration of Multi-Vendor LIS EnvironmentsDocument59 pages032-Roubort-Advantages and Integration of Multi-Vendor LIS EnvironmentsSukma EffendyNo ratings yet
- 032-Roubort-Advantages and Integration of Multi-Vendor LIS EnvironmentsDocument59 pages032-Roubort-Advantages and Integration of Multi-Vendor LIS EnvironmentsSukma EffendyNo ratings yet
- MRSA Recommendations: Members of The Task ForceDocument13 pagesMRSA Recommendations: Members of The Task ForceNasrullah Samejo 2No ratings yet
- Building A Smart Laboratory: An Introduction To The Integrated LabDocument36 pagesBuilding A Smart Laboratory: An Introduction To The Integrated LabSukma EffendyNo ratings yet
- 032-Roubort-Advantages and Integration of Multi-Vendor LIS EnvironmentsDocument59 pages032-Roubort-Advantages and Integration of Multi-Vendor LIS EnvironmentsSukma EffendyNo ratings yet
- Serial NumberDocument1 pageSerial NumberSukma EffendyNo ratings yet
- MRSA Infections: Jama Patient PageDocument1 pageMRSA Infections: Jama Patient PageSukma EffendyNo ratings yet
- 032-Roubort-Advantages and Integration of Multi-Vendor LIS EnvironmentsDocument59 pages032-Roubort-Advantages and Integration of Multi-Vendor LIS EnvironmentsSukma EffendyNo ratings yet
- DM Lipid Profile With LDL PDFDocument1 pageDM Lipid Profile With LDL PDFSukma EffendyNo ratings yet
- An Overview of The Pulmonary SystemDocument9 pagesAn Overview of The Pulmonary SystemSukma EffendyNo ratings yet
- 02 RX Monaco (RX5000)Document1 page02 RX Monaco (RX5000)Sukma EffendyNo ratings yet
- Anemia MonographDocument140 pagesAnemia MonographZaveri Hemant GirishkumarNo ratings yet
- In Flight ArabicDocument12 pagesIn Flight ArabicSukma EffendyNo ratings yet
- In Flight ArabicDocument12 pagesIn Flight ArabicSukma EffendyNo ratings yet
- Study of Lipid Peroxide and Lipid Profile in Diabetes Mellitus PDFDocument5 pagesStudy of Lipid Peroxide and Lipid Profile in Diabetes Mellitus PDFSukma EffendyNo ratings yet
- DM Lipid Profile With LDL PDFDocument1 pageDM Lipid Profile With LDL PDFSukma EffendyNo ratings yet
- A Study of Oxidative Stress, Antioxidant Status and Lipid Profile in Diabetic Patient in The Western Region of Nepal PDFDocument7 pagesA Study of Oxidative Stress, Antioxidant Status and Lipid Profile in Diabetic Patient in The Western Region of Nepal PDFSukma EffendyNo ratings yet
- National Geographic 2010-05Document164 pagesNational Geographic 2010-05arsivlik12100% (1)
- Surveys Manual Glossary PDFDocument57 pagesSurveys Manual Glossary PDFSukma EffendyNo ratings yet
- Detection of VWF PDFDocument11 pagesDetection of VWF PDFSukma EffendyNo ratings yet
- Validated Sandwich ELISA For The Quantification of Von Willebrand Fact - PDF - 3236 PDFDocument9 pagesValidated Sandwich ELISA For The Quantification of Von Willebrand Fact - PDF - 3236 PDFSukma EffendyNo ratings yet
- Evaluation of The GeneXpert MTBRIF Assay For Rapid Diagnosis TB ElimDocument5 pagesEvaluation of The GeneXpert MTBRIF Assay For Rapid Diagnosis TB ElimSukma EffendyNo ratings yet
- Vascular Permeability ICAM-1: Role in Inflammation and in The Regulation ofDocument3 pagesVascular Permeability ICAM-1: Role in Inflammation and in The Regulation ofSukma EffendyNo ratings yet
- Detection of Mycobacterium Tuberculosis in Blood Using The Xpert MTBRIF AssayDocument28 pagesDetection of Mycobacterium Tuberculosis in Blood Using The Xpert MTBRIF AssaySukma EffendyNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Three Types of Connective Tissue Proper Loose Dense SpecializedDocument3 pagesThree Types of Connective Tissue Proper Loose Dense SpecializedStevNo ratings yet
- Immuno Hematology 25Document56 pagesImmuno Hematology 25BRITTO12No ratings yet
- Journal ReadingDocument18 pagesJournal ReadingmitaNo ratings yet
- Coadata 504+: Incl. Dual-Wavelength TechnologyDocument4 pagesCoadata 504+: Incl. Dual-Wavelength TechnologyAgung PutraNo ratings yet
- 1 Blood Transfusion Practice and Quality Control PDFDocument17 pages1 Blood Transfusion Practice and Quality Control PDFWenz MacuteNo ratings yet
- Tata Laksana Perawatan Gigi Pada Pasien Kelainan Jantung Yang Ditanamkan Icd Dan Mengonsumsi WarfarinDocument17 pagesTata Laksana Perawatan Gigi Pada Pasien Kelainan Jantung Yang Ditanamkan Icd Dan Mengonsumsi WarfarinEka ApriantiNo ratings yet
- Abo Blood Group SystemDocument10 pagesAbo Blood Group SystemMariel AbatayoNo ratings yet
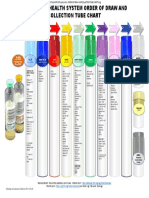
- Start Here: Gold Pink White EDTA W/ Plasma SeparatorDocument1 pageStart Here: Gold Pink White EDTA W/ Plasma SeparatorfdsfdsfNo ratings yet
- Nodes of RanvierDocument25 pagesNodes of RanvierHariharanNo ratings yet
- Evaluation of The Ruminant Complete Blood Dell CountDocument26 pagesEvaluation of The Ruminant Complete Blood Dell CountDirección Científica Laboratorio VitalabNo ratings yet
- Tablas de Presión ArterialDocument2 pagesTablas de Presión Arterialwasserpura100% (1)
- ZGMzZGE3ODAzZjU0ZjEyNjBkOTI2ZDg1YjAwZDY1OGQ2NzM4MDY5Yg PDFDocument137 pagesZGMzZGE3ODAzZjU0ZjEyNjBkOTI2ZDg1YjAwZDY1OGQ2NzM4MDY5Yg PDFdedyNo ratings yet
- Chapter 8: Blood Rheology: Christina KolyvaDocument24 pagesChapter 8: Blood Rheology: Christina Kolyvaclacsik bikin asikNo ratings yet
- Unit 2 Bio LAB2Document2 pagesUnit 2 Bio LAB2Travis JohnsonNo ratings yet
- Polycythemia Vera - Blood Disorders - MSD Manual Family Health VersionDocument4 pagesPolycythemia Vera - Blood Disorders - MSD Manual Family Health VersionShoaib BajwaNo ratings yet
- Practical HaematologyDocument70 pagesPractical HaematologyMustafa Khandgawi100% (1)
- CH 16 BloodDocument40 pagesCH 16 BloodShampa SenNo ratings yet
- Bones Joints Anatomy Week 3Document5 pagesBones Joints Anatomy Week 3hassan razaNo ratings yet
- Tissues: 1. Objective QuestionsDocument7 pagesTissues: 1. Objective Questionsnaman mahawer100% (1)
- Lab Muscle Tissue EditinggDocument12 pagesLab Muscle Tissue EditinggLyn YmiNo ratings yet
- Haematological Profile and Erythrocyte Indices in Different Breeds of PoultryDocument4 pagesHaematological Profile and Erythrocyte Indices in Different Breeds of PoultrySukma WijayaNo ratings yet
- Pengaruh Lama Penundaan Pengecatan Setelah Fiksasi Apusan Darah Tepi Terhadap Morfologi EritrositDocument5 pagesPengaruh Lama Penundaan Pengecatan Setelah Fiksasi Apusan Darah Tepi Terhadap Morfologi EritrositAlissa QothrunnadaNo ratings yet
- HEMATOLOGY Chapter 20 Rodaks Hematology, 5E (2016)Document14 pagesHEMATOLOGY Chapter 20 Rodaks Hematology, 5E (2016)Sachi Xandria de LaraNo ratings yet
- Sept 2019 Hematology NotesDocument57 pagesSept 2019 Hematology NotesDayanna NarcisoNo ratings yet
- Coagulation - LabDocument7 pagesCoagulation - LabGuia De GuzmanNo ratings yet
- Iesc 106Document15 pagesIesc 106pjatinNo ratings yet
- Rest of No 1Document16 pagesRest of No 1eman el saeedNo ratings yet
- Muscular SystemDocument36 pagesMuscular SystemDeepak KdNo ratings yet
- Tissue Anatomy and PhysiologyDocument35 pagesTissue Anatomy and Physiologysnp88100% (1)
- ABO Incompatibility Disease of The NewbornDocument10 pagesABO Incompatibility Disease of The NewbornupuanNo ratings yet