You might also like
- Reacciones químicas no isotérmicas: Ecuaciones de balance de energíaDocument50 pagesReacciones químicas no isotérmicas: Ecuaciones de balance de energíauni2004No ratings yet
- Criterio de Equilibrio Sistemas Con Reacciones SimplesDocument18 pagesCriterio de Equilibrio Sistemas Con Reacciones SimplesMario FloresNo ratings yet
- Termoquímica: Calores de reacción, entalpía y entropíaDocument44 pagesTermoquímica: Calores de reacción, entalpía y entropíaCarolina Tejada GomezNo ratings yet
- Problema de TermodinámicaDocument52 pagesProblema de TermodinámicaDonek SantanderNo ratings yet
- Unidad III Energía Libre y Equilibrio, FQ II Sem. 2020BDocument20 pagesUnidad III Energía Libre y Equilibrio, FQ II Sem. 2020Bantonio loraNo ratings yet
- REACCIONES QUÍMICAS EN FASE GASEOSA: EQUILIBRIO Y FACTORES QUE LO AFECTANDocument13 pagesREACCIONES QUÍMICAS EN FASE GASEOSA: EQUILIBRIO Y FACTORES QUE LO AFECTANMario Flores100% (1)
- Formulario de FisicoquímicaDocument2 pagesFormulario de FisicoquímicaMichel Gutierrez Silva100% (1)
- Desarrollo de FormulasDocument6 pagesDesarrollo de FormulasRichard Gonzalo YtoNo ratings yet
- Apuntes de FugacidadDocument14 pagesApuntes de FugacidadNaferMrtnzMNo ratings yet
- Propiedades Molales ParcialesDocument43 pagesPropiedades Molales ParcialesPablo Millan0% (1)
- Fórmulas TermodinámicaDocument3 pagesFórmulas TermodinámicaThiago OsorioNo ratings yet
- Notas Termodinamica Del EquilibrioDocument94 pagesNotas Termodinamica Del EquilibrioIsseiNo ratings yet
- Ej Fugacidad MezclasDocument9 pagesEj Fugacidad MezclasAndrés NorambuenaNo ratings yet
- Taller de Fisico2Document4 pagesTaller de Fisico2JUAN MIGUEL BEDOYA MIRANDANo ratings yet
- Fi2004 CfasesDocument25 pagesFi2004 CfasesAurelioMarco007No ratings yet
- Revision de Termodinamica Fisico Quimica-Me211rDocument35 pagesRevision de Termodinamica Fisico Quimica-Me211rCarlos Catalán CórdovaNo ratings yet
- Ecuación de estado gas ideal PV=nRTDocument12 pagesEcuación de estado gas ideal PV=nRTJose Alonzo VivancosNo ratings yet
- Energía Libre y EquilibrioDocument15 pagesEnergía Libre y EquilibrioEloisa Amell AmellNo ratings yet
- Ter Cera LeyDocument8 pagesTer Cera LeyVllsSNo ratings yet
- Constante de Equilibrio QuimicoDocument8 pagesConstante de Equilibrio QuimicoCamila Natalia Zapata AlvaradoNo ratings yet
- Ingeniería Electromecánica.: Máquinas Y Equipos Térmicos 1. Complemento de La Unidad 1Document13 pagesIngeniería Electromecánica.: Máquinas Y Equipos Térmicos 1. Complemento de La Unidad 1Kevin PerezNo ratings yet
- Balanceenergia 10356Document22 pagesBalanceenergia 10356Jugo AmenNo ratings yet
- Diseño de Reactores: Efectos de Temperatura y PresiónDocument12 pagesDiseño de Reactores: Efectos de Temperatura y PresiónJosefaNo ratings yet
- ENTROPIADocument5 pagesENTROPIAJhonatan Meza AparicioNo ratings yet
- Segundo Principio TermodinamicoDocument12 pagesSegundo Principio TermodinamicoJose Severiano Luis Bravo Mora. Ph.D.No ratings yet
- Constante de FugacidadDocument9 pagesConstante de FugacidadSara CristinaNo ratings yet
- Resumen Reactores QuímicosDocument20 pagesResumen Reactores QuímicosSara Mesa MedinaNo ratings yet
- Energias Libres Problemas ResueltosDocument10 pagesEnergias Libres Problemas ResueltosArturo Barradas NNo ratings yet
- Ejercicios Física AplicadaDocument6 pagesEjercicios Física AplicadaNATALIA ORTEGA GOMEZNo ratings yet
- Termodinamica - Modulo 3 - PARTE 1Document37 pagesTermodinamica - Modulo 3 - PARTE 1Ety Viloria CastilloNo ratings yet
- Taller 4 FQ 2022-1Document5 pagesTaller 4 FQ 2022-1humberto lopezNo ratings yet
- Unidad 1Document45 pagesUnidad 1n_a_n_i_t_aNo ratings yet
- Isoterma de ReaccionDocument13 pagesIsoterma de ReaccionAlma Sosa FloresNo ratings yet
- Apuntes E. Libre y Eq. Fís. Sust. Puras 2021BDocument5 pagesApuntes E. Libre y Eq. Fís. Sust. Puras 2021BBrenda Lizbeth Guillen SustaitaNo ratings yet
- Gases Chapi 1Document6 pagesGases Chapi 1reynaldoNo ratings yet
- Chuletario de Termodinámica.Document3 pagesChuletario de Termodinámica.ap0cal1pse100% (1)
- SESION 9-Termoquìmica.Document29 pagesSESION 9-Termoquìmica.Gabriel Armando Orellana NolascoNo ratings yet
- Desarrollo de FormulasDocument4 pagesDesarrollo de FormulasRichard Gonzalo YtoNo ratings yet
- Leyes de los gases idealesDocument9 pagesLeyes de los gases idealesberenice serrano torresNo ratings yet
- Estimación y cálculo de fugacidades en mezclas de componentesDocument5 pagesEstimación y cálculo de fugacidades en mezclas de componentesGonzalo vargasNo ratings yet
- Informe 3Document12 pagesInforme 3Jose Miguel TrujilloNo ratings yet
- QUIMICA FISICA II - Equilibrios de fases y transicionesDocument52 pagesQUIMICA FISICA II - Equilibrios de fases y transicionesJaviera Paz ValenzuelaNo ratings yet
- Equilibrio Quimico y Equilibrio FisicoDocument34 pagesEquilibrio Quimico y Equilibrio FisicoJussef Fuentes QuinteroNo ratings yet
- Capitulo1 (Gases)Document34 pagesCapitulo1 (Gases)Jos Man Quisbert EspinozaNo ratings yet
- Termodinámica Las SolucionesDocument21 pagesTermodinámica Las SolucionesEl Angel CortezNo ratings yet
- 3a - ENERGÍA LIBRE DE GIBBS Y EQUILIBRIO QUÍMICODocument11 pages3a - ENERGÍA LIBRE DE GIBBS Y EQUILIBRIO QUÍMICOMaximiliano LorenzanaNo ratings yet
- Microsoft Word - Res - TermoquimicaDocument2 pagesMicrosoft Word - Res - TermoquimicaÀlex Ripoll CollNo ratings yet
- Transiciones de Fase y Estados Criticos (Resumen)Document10 pagesTransiciones de Fase y Estados Criticos (Resumen)Erick Ntl TlvNo ratings yet
- Temperatura aproximada 363,35 KDocument5 pagesTemperatura aproximada 363,35 KJessenia CruzNo ratings yet
- Interaccion TermicaDocument48 pagesInteraccion TermicajagrlzzgNo ratings yet
- Formulario FISICOQUIMICADocument2 pagesFormulario FISICOQUIMICAquiron001100% (1)
- APLICACIONES-1 de Ecuaciones DiferencialesDocument14 pagesAPLICACIONES-1 de Ecuaciones DiferencialesLuis Pablo Ramírez RomeroNo ratings yet
- Equilibrio de reacciones con fases condensadas y gaseosasDocument30 pagesEquilibrio de reacciones con fases condensadas y gaseosasAlexander TRNo ratings yet
- Segunda, Tercera Ley y Energía LibreDocument18 pagesSegunda, Tercera Ley y Energía LibreCESAR ANTONIO DIAZ CISNEROSNo ratings yet
- Ingeniería química. Soluciones a los problemas del tomo IFrom EverandIngeniería química. Soluciones a los problemas del tomo INo ratings yet
- Examen Matemáticas II de Asturias (Extraordinaria de 1998) (WWW - Examenesdepau.com)Document1 pageExamen Matemáticas II de Asturias (Extraordinaria de 1998) (WWW - Examenesdepau.com)Juan García MenéndezNo ratings yet
- Atlas Nacional de España. Sección II. Climatología 2005 2edDocument36 pagesAtlas Nacional de España. Sección II. Climatología 2005 2edJuan García MenéndezNo ratings yet
- CASTILLA Y LEON Septiembre 2010 G PDFDocument2 pagesCASTILLA Y LEON Septiembre 2010 G PDFPianistasenderistaNo ratings yet
- Examen Física de Asturias (Ordinaria de 1998) (WWW - Examenesdepau.com)Document1 pageExamen Física de Asturias (Ordinaria de 1998) (WWW - Examenesdepau.com)Juan García MenéndezNo ratings yet
- Teléfonos Operativos LEIB - v25Document1 pageTeléfonos Operativos LEIB - v25Juan García MenéndezNo ratings yet
- 2018 11 06 Cantabria Report - RT - 0455 - 2018Document4 pages2018 11 06 Cantabria Report - RT - 0455 - 2018Juan García MenéndezNo ratings yet
- Hidrología 2007 2edDocument56 pagesHidrología 2007 2edJuan García MenéndezNo ratings yet
- 2019 06 10 Cantabria Report - RT - 0207 - 2019Document7 pages2019 06 10 Cantabria Report - RT - 0207 - 2019Juan García MenéndezNo ratings yet
- Física Del Caos en La Predicción MeteorológicaDocument1,198 pagesFísica Del Caos en La Predicción MeteorológicaJuan García MenéndezNo ratings yet
- 2018-11-06-Asturias-Notificacion de Resolucion de acceso-sindatos-SinFirmasTrasNotificacion23ene2020Document4 pages2018-11-06-Asturias-Notificacion de Resolucion de acceso-sindatos-SinFirmasTrasNotificacion23ene2020Juan García MenéndezNo ratings yet
- Adenda Electromagnetismo USDocument4 pagesAdenda Electromagnetismo USJuan García MenéndezNo ratings yet
- 2018 11 06 Cantabria Report - RT - 0455 - 2018Document4 pages2018 11 06 Cantabria Report - RT - 0455 - 2018Juan García MenéndezNo ratings yet
- Mecánica de Fluidos by Amable Liñán Martínez, M. Rodríguez Fernández, F.J. Higuera AntónDocument193 pagesMecánica de Fluidos by Amable Liñán Martínez, M. Rodríguez Fernández, F.J. Higuera AntónJuan García Menéndez50% (2)
- 2019-02-04-Acord Resol Exp. 42-13326-18-SinFirmasTrasNotificacion23ene2020Document10 pages2019-02-04-Acord Resol Exp. 42-13326-18-SinFirmasTrasNotificacion23ene2020Juan García MenéndezNo ratings yet
- NotasAutobiograficasEinstein PDFDocument37 pagesNotasAutobiograficasEinstein PDFJuan García MenéndezNo ratings yet
- IndexDocument224 pagesIndexmarco100% (1)
- 2018-10-05-Asturias-García Simón Enrique - traslado-sindatos-SinFirmasTrasNotificacion23ene2020Document1 page2018-10-05-Asturias-García Simón Enrique - traslado-sindatos-SinFirmasTrasNotificacion23ene2020Juan García MenéndezNo ratings yet
- Analisis Numerico Primer Curso Hernan GonzalezDocument195 pagesAnalisis Numerico Primer Curso Hernan Gonzalezcarjuan18100% (3)
- FM Javier MasDocument150 pagesFM Javier MasJuan Negredo GarciaNo ratings yet
- 12lhilbert PDFDocument35 pages12lhilbert PDFAdrian Pavon DuarteNo ratings yet
- Javier BUCETA FERNÁNDEZ - Elka Radoslavova KOROUTCHEVA - Juan Manuel PASTOR RUIZ - Temas de Biofísica (2012, Editorial UNED)Document340 pagesJavier BUCETA FERNÁNDEZ - Elka Radoslavova KOROUTCHEVA - Juan Manuel PASTOR RUIZ - Temas de Biofísica (2012, Editorial UNED)Andrés Lopez100% (4)
- Física de PartículasDocument102 pagesFísica de Partículasasdfg_100% (3)
- Introduccion A La Mecanica Cuantica PDFDocument276 pagesIntroduccion A La Mecanica Cuantica PDFJhon F. RodriguezNo ratings yet
- Física Nuclear y de Partículas - Problemas Resueltos PDFDocument259 pagesFísica Nuclear y de Partículas - Problemas Resueltos PDFJuan García Menéndez83% (6)
- Vademecum PDFDocument308 pagesVademecum PDFRodrigo Bravo EspejoNo ratings yet
- Int Fisica Cuantica-RetamosaDocument231 pagesInt Fisica Cuantica-Retamosaelke_hkNo ratings yet
- 100 Problemas de Fisica Cuantica PDFDocument308 pages100 Problemas de Fisica Cuantica PDFmushu27100% (1)
- Introduccion A La Fisica CuanticaDocument225 pagesIntroduccion A La Fisica CuanticaDonax Sanchez CardenasNo ratings yet
- UNED. Química Cuántica - La Química Cuántica en 100 Problemas - Lorna Elizabeth Bailey Chapman, María Dolores Troitiño NúñezDocument234 pagesUNED. Química Cuántica - La Química Cuántica en 100 Problemas - Lorna Elizabeth Bailey Chapman, María Dolores Troitiño NúñezJuan García Menéndez100% (5)
- Interferencia OpticaDocument37 pagesInterferencia OpticaJuan García MenéndezNo ratings yet
- Relleno Doña JuanaDocument13 pagesRelleno Doña JuanaEsteban AdarveNo ratings yet
- Documento Guia 6 CienciasDocument2 pagesDocumento Guia 6 CienciasJose angel Marquez oviedoNo ratings yet
- Ensayo XDDocument12 pagesEnsayo XDJuan David Martinez VargasNo ratings yet
- LBFQ Clase2Document22 pagesLBFQ Clase2Branco Palma RodriguezNo ratings yet
- Procesos Industriales Ii: Mg. Pedro Arias Cubillas Ilo, 2022Document24 pagesProcesos Industriales Ii: Mg. Pedro Arias Cubillas Ilo, 2022xiory estefanyNo ratings yet
- PolimetrosDocument15 pagesPolimetrosJessy MontoyaNo ratings yet
- Caolin PectinaDocument2 pagesCaolin PectinaJavan Zaldivar100% (1)
- Viroc CarpetaTecnicaDocument29 pagesViroc CarpetaTecnicaOctavio CuevasNo ratings yet
- Fibras Dietarias y Su Aplicación en El Desarrollo PDFDocument97 pagesFibras Dietarias y Su Aplicación en El Desarrollo PDFMarco Montoya JuradoNo ratings yet
- Amoniaco AcidonitricoDocument15 pagesAmoniaco AcidonitricoDanny AndradeNo ratings yet
- Calentamiento y Climatización de PiscinasDocument7 pagesCalentamiento y Climatización de PiscinasMiguel de la torreNo ratings yet
- Manometros WIKADocument3 pagesManometros WIKAGabriel DittamoNo ratings yet
- Investigacion Seminario 3Document10 pagesInvestigacion Seminario 3LuceroNo ratings yet
- Guía LFQII (EPIQ) 2021-2Document38 pagesGuía LFQII (EPIQ) 2021-2Carlos Jesús Fabián ArévaloNo ratings yet
- Celdas de FlotacionDocument6 pagesCeldas de FlotacionLuis PalmaNo ratings yet
- Problemas Resueltos de La Lección 6Document31 pagesProblemas Resueltos de La Lección 6Ayde Rosario Alconz IngalaNo ratings yet
- Preinforme Caida de Presion Con Accesorios (FDocument9 pagesPreinforme Caida de Presion Con Accesorios (F0231720008 ODAIR MENDOZA CABARCAS ESTUDIANTE ACTIVONo ratings yet
- Exa 3 QM1 Julio 2022Document1 pageExa 3 QM1 Julio 2022marcelo silvera moliNo ratings yet
- Ficha Técnica BRASH 2021Document1 pageFicha Técnica BRASH 2021Abraham Martínez GómezNo ratings yet
- Clasificación de Los MaterialesDocument8 pagesClasificación de Los MaterialesProject ElizabethNo ratings yet
- Shell Tactic Customer PresentationDocument50 pagesShell Tactic Customer Presentationfreddy william montalban palaciosNo ratings yet
- Laboratorio de Plomo MIERCOLES (8-12)Document9 pagesLaboratorio de Plomo MIERCOLES (8-12)Edson RiveraNo ratings yet
- Lab Ciclo de RefrigeraciónDocument7 pagesLab Ciclo de Refrigeraciónmaria gonzalezNo ratings yet
- Clase #7 - PtapDocument16 pagesClase #7 - PtapGRETTY EDITH TAPARA TUNCARNo ratings yet
- Los PlásticosDocument17 pagesLos PlásticosJulia Cmon100% (1)
- Secado Directo A Condiciones ConstantesDocument34 pagesSecado Directo A Condiciones ConstantesPamela VargasNo ratings yet
- TAMIZAJE FITOQUIMICO DE HOJAS Y SEMILLAS DE (Bixa Orellana) - TANIA GUERRERO VEJARANO, - JOSE LUIS PAREDES SALAZARDocument15 pagesTAMIZAJE FITOQUIMICO DE HOJAS Y SEMILLAS DE (Bixa Orellana) - TANIA GUERRERO VEJARANO, - JOSE LUIS PAREDES SALAZAREver BerriosNo ratings yet
- Práctica de TermometríaDocument2 pagesPráctica de TermometríaMarlon VásquezNo ratings yet
- Apu Estructuras Metalicas VigasDocument3 pagesApu Estructuras Metalicas VigasJdcad IngenierosNo ratings yet
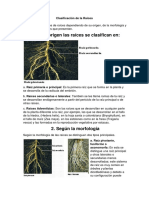
- Clasificación de La RaicesDocument7 pagesClasificación de La RaicesExcel Marroquin G.No ratings yet