You might also like
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Slot 22-23-24 Real Time CommunicationDocument50 pagesSlot 22-23-24 Real Time CommunicationLinh BuiNo ratings yet
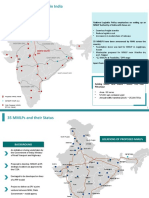
- Development of Mmlps in India: Western DFC Eastern DFCDocument2 pagesDevelopment of Mmlps in India: Western DFC Eastern DFCsdfg100% (2)
- Role of Optimizer in Neural NetworkDocument2 pagesRole of Optimizer in Neural NetworkMuhammad AlianNo ratings yet
- PE1 Introduction To MovementsDocument4 pagesPE1 Introduction To MovementsDaniela AnsayNo ratings yet
- 1663-43691010-Me-Dwg-7380-00-Cu-Cl2-Bdu-Fru-Assy-1 (R1)Document1 page1663-43691010-Me-Dwg-7380-00-Cu-Cl2-Bdu-Fru-Assy-1 (R1)raacontreras21No ratings yet
- World Is Flat Thomas FriedmanDocument10 pagesWorld Is Flat Thomas FriedmanGilean DalidaNo ratings yet
- So You Want To Have Your Own Cosmetics BrandDocument3 pagesSo You Want To Have Your Own Cosmetics BrandCarlosNo ratings yet
- Yuasa Technical Data Sheet: The World's Leading Battery ManufacturerDocument1 pageYuasa Technical Data Sheet: The World's Leading Battery ManufacturerAshraf Sayed ShabaanNo ratings yet
- Ai R16 - Unit-6Document36 pagesAi R16 - Unit-6RakeshNo ratings yet
- FOCGB5 Rtest 1-6BDocument3 pagesFOCGB5 Rtest 1-6BM-DNo ratings yet
- University of Central Punjab: Object Oriented ProgrammingDocument3 pagesUniversity of Central Punjab: Object Oriented ProgrammingChoudhary MuneebNo ratings yet
- Land SurveyingDocument174 pagesLand SurveyingArifulmakrif Hailee100% (2)
- Module 2: Most Essential Learning Competencies (Melcs) : Lesson 1: Background, Rationale, and Development of MelcsDocument5 pagesModule 2: Most Essential Learning Competencies (Melcs) : Lesson 1: Background, Rationale, and Development of MelcsRobert Kier Tanquerido TomaroNo ratings yet
- Optimal Control Development System For ElectricalDocument7 pagesOptimal Control Development System For ElectricalCRISTIAN CAMILO MORALES SOLISNo ratings yet
- Rae Systems Parts List October 26 2010Document2 pagesRae Systems Parts List October 26 2010Mike HastingsNo ratings yet
- Technical Bulletin: ConditionDocument11 pagesTechnical Bulletin: ConditionFrancoNo ratings yet
- Links of XII Physics Investigatory Project Allowtment 2Document3 pagesLinks of XII Physics Investigatory Project Allowtment 2yashtomarsoop07No ratings yet
- Missouri Department of Transportation: Standard Inspection and Testing Plan (ITP)Document32 pagesMissouri Department of Transportation: Standard Inspection and Testing Plan (ITP)mohamedamine.zemouriNo ratings yet
- Term Paper On Strategic ManagementDocument8 pagesTerm Paper On Strategic Managementea7j5ys3100% (1)
- Audio Level Meter - ProjectDocument4 pagesAudio Level Meter - ProjectMircea PanzariuNo ratings yet
- DAC AnalysisDocument19 pagesDAC Analysisమురళీధర్ ఆది ఆంధ్రుడుNo ratings yet
- D6588Document6 pagesD6588Pulak DebnathNo ratings yet
- Classical Electromagnetism 1st Edition Franklin Solutions ManualDocument21 pagesClassical Electromagnetism 1st Edition Franklin Solutions ManualBrianYorktnqsw100% (15)
- 17333Document2 pages17333Nithya RaviNo ratings yet
- Gerrard 1966Document13 pagesGerrard 1966AnandhuMANo ratings yet
- Reading TAF'sDocument4 pagesReading TAF'sDouglas AlvarezNo ratings yet
- Dubai Holding FactsheetDocument2 pagesDubai Holding FactsheetHarley soulNo ratings yet
- Module 4 - Community Health AssessmentDocument8 pagesModule 4 - Community Health AssessmentSteffiNo ratings yet
- Daftar Isian 3 Number Plate, Danger Plate, Anti Climbing DeviceDocument2 pagesDaftar Isian 3 Number Plate, Danger Plate, Anti Climbing DeviceMochammad Fauzian RafsyanzaniNo ratings yet
- 3PM - Project Management Method - Sales MessagingDocument5 pages3PM - Project Management Method - Sales Messagingberuang kutubNo ratings yet