You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Noun Worksheet For Class 4 With Answers 1Document10 pagesNoun Worksheet For Class 4 With Answers 1Kamini jain100% (3)
- Level 2 Diploma Health and Social Care: Unit 22 - Undertake Agreed Pressure Area CareDocument6 pagesLevel 2 Diploma Health and Social Care: Unit 22 - Undertake Agreed Pressure Area CareJayne AshleyNo ratings yet
- A Study of The Atetela PeopleDocument60 pagesA Study of The Atetela Peoplewelovell100% (2)
- Alphabetical List of English Proverbs - L-ZDocument5 pagesAlphabetical List of English Proverbs - L-ZHendra TaufikNo ratings yet
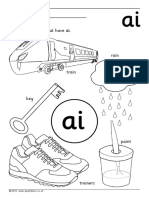
- Jolly PhonicsDocument22 pagesJolly PhonicsJack Murphy100% (2)
- Plants and Animals of The Book of MormonDocument42 pagesPlants and Animals of The Book of MormonMosiah221100% (3)
- F Y) Y, 0 y 3: Fuad Adrian Iskandar 1606843722 PR-I Matematika Terapan (Statistik)Document2 pagesF Y) Y, 0 y 3: Fuad Adrian Iskandar 1606843722 PR-I Matematika Terapan (Statistik)Nadia KarimaNo ratings yet
- LGBTQ Z?: Hypatia Vol. 27, No. 3 (Summer 2012) © by Hypatia, IncDocument15 pagesLGBTQ Z?: Hypatia Vol. 27, No. 3 (Summer 2012) © by Hypatia, IncSouthern PrideNo ratings yet
- Canadian Geographic Best Wildlife Photography 2018Document101 pagesCanadian Geographic Best Wildlife Photography 2018glezglez1No ratings yet
- Anchorage Control in Bioprogressive Vs Straight-Wire Treatment PDFDocument6 pagesAnchorage Control in Bioprogressive Vs Straight-Wire Treatment PDFsolodont1No ratings yet
- George WashingtonDocument12 pagesGeorge Washingtonsubathra samiveluNo ratings yet
- Bikalpa 19Document28 pagesBikalpa 19Ebang BikalpaNo ratings yet
- The Shameful Exploits of Rinku BhabhiDocument91 pagesThe Shameful Exploits of Rinku BhabhiSriram Kumar0% (1)
- Case Taking FormatDocument29 pagesCase Taking FormatmadhukarNo ratings yet
- Factors Influencing ToxicityDocument7 pagesFactors Influencing ToxicityderrickNo ratings yet
- Pourusha VastiDocument54 pagesPourusha Vastiksr prasadNo ratings yet
- Pengetahuan Dan Pemahaman Umum 13 - 20 PDFDocument7 pagesPengetahuan Dan Pemahaman Umum 13 - 20 PDFMohamad Faruq ImaduddinNo ratings yet
- Animal Idioms: It'S A Jungle Out There!Document20 pagesAnimal Idioms: It'S A Jungle Out There!Edgar Bonilla MuñozNo ratings yet
- Paper 2 Biology 1995 Paper 2+ansDocument30 pagesPaper 2 Biology 1995 Paper 2+ansapi-3812894No ratings yet
- Kasus Myasis Yang Disebabkan Oleh Chrysomya Bezziana Di Pulau JawaDocument8 pagesKasus Myasis Yang Disebabkan Oleh Chrysomya Bezziana Di Pulau JawaIswandiIbnAmirNo ratings yet
- Animals HomesDocument6 pagesAnimals HomesFlor Stan50% (2)
- CLINIC CASE Histoplasmosis SoluciónDocument2 pagesCLINIC CASE Histoplasmosis SoluciónCarolina Mueses0% (1)
- Cleft and PalateDocument77 pagesCleft and PalateNini BendelianiNo ratings yet
- Facts About Owls: Horned Owl (Bubo Virginianus, Fig. 1) Is The Most Widely Distributed Owl inDocument6 pagesFacts About Owls: Horned Owl (Bubo Virginianus, Fig. 1) Is The Most Widely Distributed Owl insprite2lNo ratings yet
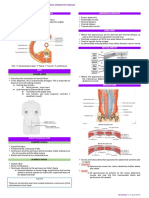
- Ob - Maternal AnatomyDocument8 pagesOb - Maternal AnatomyLeslee Cruz - BernardinoNo ratings yet
- Keys For Pet Exam PackDocument34 pagesKeys For Pet Exam PackEleftheria AggeleaNo ratings yet
- Singular Answer KeyDocument2 pagesSingular Answer KeyEuclid Mark83% (6)
- What Is Ovarian CancerDocument2 pagesWhat Is Ovarian CancerArianaNo ratings yet
- Class 11 Biology NCERT Textbook Chapter 1 The Living WorldDocument20 pagesClass 11 Biology NCERT Textbook Chapter 1 The Living WorldROHAN SHARMANo ratings yet