You might also like
- Spiders and Thier Kin - Golden Guide 1990 PDFDocument164 pagesSpiders and Thier Kin - Golden Guide 1990 PDFConifor100% (1)
- Cranial NerveDocument39 pagesCranial NerveСуджива Гунасинха100% (2)
- Neuron and Glial CellsDocument8 pagesNeuron and Glial CellsMurilo Bonet100% (1)
- Garuda PuranaDocument8 pagesGaruda PuranaSourav RanjanNo ratings yet
- HIstology Experiment 12Document5 pagesHIstology Experiment 12Pearlregine Cianne MirandaNo ratings yet
- Anatomy of RetinaDocument39 pagesAnatomy of Retinadr_dev100% (1)
- Excitable Cells: Monographs in Modern Biology for Upper School and University CoursesFrom EverandExcitable Cells: Monographs in Modern Biology for Upper School and University CoursesNo ratings yet
- Animal Behavior Chapter 1Document14 pagesAnimal Behavior Chapter 1StudentNo ratings yet
- Anatomy of Eye: By: Dr. Lubna ShahDocument36 pagesAnatomy of Eye: By: Dr. Lubna ShahRida ShahNo ratings yet
- Anatomy and Physiology of The Human EyeDocument9 pagesAnatomy and Physiology of The Human EyeMaryy Basilio100% (2)
- Histology - Nerve Tissue and The Nervous SystemDocument21 pagesHistology - Nerve Tissue and The Nervous SystemDan UvarovNo ratings yet
- Simple Anatomy of The RetinaDocument24 pagesSimple Anatomy of The RetinaLateepheart YakubNo ratings yet
- Visual SystemDocument56 pagesVisual SystemMartha MulusaNo ratings yet
- For The Electronics Screen, See - For Other Uses, See .: Jump To Navigation Jump To SearchDocument10 pagesFor The Electronics Screen, See - For Other Uses, See .: Jump To Navigation Jump To SearchNicoleta TuleaNo ratings yet
- Retina Anatomy - Overview, Gross Anatomy, Microscopic AnatomyDocument3 pagesRetina Anatomy - Overview, Gross Anatomy, Microscopic AnatomyjackNo ratings yet
- Sreejith TDocument37 pagesSreejith TAbdisamed AllaaleNo ratings yet
- Histology of Cornea & RetinaDocument14 pagesHistology of Cornea & RetinaSandhya KurupNo ratings yet
- Neuroanatomy, Pupillary Light Reflexes and PathwayDocument6 pagesNeuroanatomy, Pupillary Light Reflexes and PathwayMatthew PrestonNo ratings yet
- Bionic Eye EB KNS 2019Document58 pagesBionic Eye EB KNS 2019Lalitaditya DivakarlaNo ratings yet
- Macroscopic Neuroanatomy: Neuroanatomy Deals With The Structure of TheDocument14 pagesMacroscopic Neuroanatomy: Neuroanatomy Deals With The Structure of TheRoziel Anne BasaNo ratings yet
- Lo Week 3 Blok Sss - April 2Document12 pagesLo Week 3 Blok Sss - April 2mellyNo ratings yet
- Hassan Nawaz - 202321192010Document15 pagesHassan Nawaz - 202321192010hassanNo ratings yet
- Fundamental72-75 in EnglishDocument5 pagesFundamental72-75 in EnglishMonika AyuningrumNo ratings yet
- Salazar - 2019 IntechOpen - Anatomy of The Human Optic Nerve - Structure and FuncionDocument46 pagesSalazar - 2019 IntechOpen - Anatomy of The Human Optic Nerve - Structure and FuncionLee Vo AnNo ratings yet
- Anatomy of The Human Optic Nerve: Structure and Function: Provisional ChapterDocument20 pagesAnatomy of The Human Optic Nerve: Structure and Function: Provisional ChapterAndres BenavidesNo ratings yet
- Anatomy of The Human Optic Nerve - Structure and FuncionDocument46 pagesAnatomy of The Human Optic Nerve - Structure and FuncionHesti hasanNo ratings yet
- Visual Pathway 2021Document83 pagesVisual Pathway 2021Pavan chowdaryNo ratings yet
- Dme C1Document7 pagesDme C1taliya. shvetzNo ratings yet
- Eye and VisionDocument19 pagesEye and VisionVincent PolicarNo ratings yet
- Anatomy of Eyes-BiopsyDocument20 pagesAnatomy of Eyes-BiopsynandanitriadiningsihNo ratings yet
- BLOK 1.3 Jaras PenglihatanDocument43 pagesBLOK 1.3 Jaras PenglihatanlenypsNo ratings yet
- The Retina: General Organization of The OrbitDocument21 pagesThe Retina: General Organization of The OrbitWendy SNo ratings yet
- EyeballDocument14 pagesEyeballMirko BelanNo ratings yet
- The Eye Pre-Lab: s94 s98 Index of ImagesDocument13 pagesThe Eye Pre-Lab: s94 s98 Index of ImagesAndi KasiantoNo ratings yet
- Sense OrgansDocument141 pagesSense OrgansitsmegkyNo ratings yet
- d8c Cranial NervesDocument29 pagesd8c Cranial NervesYhemmy Lhawal JummyNo ratings yet
- Unit 13 Sence OrganDocument132 pagesUnit 13 Sence OrganChandan ShahNo ratings yet
- Special Senses EYEDocument27 pagesSpecial Senses EYENadiabloshii NadiabloshiiNo ratings yet
- Human EyeDocument54 pagesHuman EyeRadu VisanNo ratings yet
- Embriologi Brain and SpineDocument18 pagesEmbriologi Brain and SpineJunus EufrataNo ratings yet
- Anatomy of Retina: by DR - Ashok Kumar ValuroutuDocument49 pagesAnatomy of Retina: by DR - Ashok Kumar ValuroutuanamariaboariuNo ratings yet
- Jurnal Anatomy of The Human Visual PathwayDocument7 pagesJurnal Anatomy of The Human Visual Pathwayhenry balanNo ratings yet
- ANATO - Campos VisuaisDocument11 pagesANATO - Campos VisuaishelbertpalmieroNo ratings yet
- Histology of The EyeDocument91 pagesHistology of The EyeFazira EkmaNo ratings yet
- Human Eye: A Human Eyeball Is Like A Simple Camera!Document18 pagesHuman Eye: A Human Eyeball Is Like A Simple Camera!fidansekizNo ratings yet
- Neurostiinte - Eye, Brain, Vision - David Hubel (Cap 3,4 p32-83) PDFDocument52 pagesNeurostiinte - Eye, Brain, Vision - David Hubel (Cap 3,4 p32-83) PDFgicutaNo ratings yet
- Ophthalmology 2Document235 pagesOphthalmology 2yididiyagirma30No ratings yet
- Neural Control and Coordination Grade 11Document30 pagesNeural Control and Coordination Grade 11Dr. Remya RanjithNo ratings yet
- Referat MataDocument38 pagesReferat MataAyu KottenNo ratings yet
- Animal VertebrateDocument10 pagesAnimal VertebrateAbdullahi AwwalNo ratings yet
- UntitledDocument45 pagesUntitledc/risaaq yuusuf ColoowNo ratings yet
- Development of The Central Nervous System: Consists of The Brain and Spinal Cord Derived From The Neural TubeDocument42 pagesDevelopment of The Central Nervous System: Consists of The Brain and Spinal Cord Derived From The Neural TubeZyra PerolNo ratings yet
- The Primary Visual CortexDocument30 pagesThe Primary Visual CortexΕιρήνηΔασκιωτάκηNo ratings yet
- Eye and Ear HistologyDocument28 pagesEye and Ear Histologyirakozehugue14No ratings yet
- 21 Neural Control N Coordination-NCERT Questions N AnswersDocument6 pages21 Neural Control N Coordination-NCERT Questions N AnswersGuru SrinivaasNo ratings yet
- Anatomy of The Eye and OrbitDocument7 pagesAnatomy of The Eye and OrbitmmadomadNo ratings yet
- Notes On Eye of RabbitDocument3 pagesNotes On Eye of RabbitShailesh NiroulaNo ratings yet
- Nervous Tissue: Prof. Dr. Anas Sarwar QureshiDocument12 pagesNervous Tissue: Prof. Dr. Anas Sarwar QureshiShafaqat Ghani Shafaqat GhaniNo ratings yet
- Human EyeDocument12 pagesHuman Eyeanu rettiNo ratings yet
- The EyeDocument6 pagesThe EyeDebiprasad GhoshNo ratings yet
- 1 My Trigeminal NerveDocument23 pages1 My Trigeminal NerveDan 04No ratings yet
- BMB 413 Neuroscience Online Notes Vol4Document19 pagesBMB 413 Neuroscience Online Notes Vol4Moose KeNo ratings yet
- The Cathode Ray Tube (CRT)Document5 pagesThe Cathode Ray Tube (CRT)Sourav RanjanNo ratings yet
- Heart Failure (HF), Often Referred To As Congestive Heart Failure (CHF), Occurs When TheDocument6 pagesHeart Failure (HF), Often Referred To As Congestive Heart Failure (CHF), Occurs When TheSourav RanjanNo ratings yet
- Psoriasis Is A Long-Lasting: Psoriatic Plaque, Showing A Silvery Center Surrounded by A Reddened BorderDocument2 pagesPsoriasis Is A Long-Lasting: Psoriatic Plaque, Showing A Silvery Center Surrounded by A Reddened BorderSourav RanjanNo ratings yet
- Structure: Right Atrium Right Ventricle Back Flow BloodDocument2 pagesStructure: Right Atrium Right Ventricle Back Flow BloodSourav RanjanNo ratings yet
- PlasmaDocument4 pagesPlasmaSourav RanjanNo ratings yet
- Microscopic Description: Solid Is One ofDocument5 pagesMicroscopic Description: Solid Is One ofSourav RanjanNo ratings yet
- Gas Is One ofDocument8 pagesGas Is One ofSourav RanjanNo ratings yet
- Rama (: Rāma) or Ramachandra Is TheDocument11 pagesRama (: Rāma) or Ramachandra Is TheSourav RanjanNo ratings yet
- Vishnu (: Sanskrit PronunciationDocument7 pagesVishnu (: Sanskrit PronunciationSourav RanjanNo ratings yet
- Indo-Malayan Stingless BeesDocument32 pagesIndo-Malayan Stingless BeesAbu Hassan JalilNo ratings yet
- Jurnal PoliketidaDocument43 pagesJurnal PoliketidaRama HidayatNo ratings yet
- AAZP - Case StudyDocument7 pagesAAZP - Case StudyAnil Vishwakarma0% (1)
- Animal TaxonomyDocument3 pagesAnimal TaxonomyJudy Flores100% (1)
- Z14 - Characteristics - Phylum Porifera & CnidariaDocument2 pagesZ14 - Characteristics - Phylum Porifera & CnidariaMilvy Faith C. SyjongtianNo ratings yet
- EntomologyDocument50 pagesEntomologyapi-19960301100% (1)
- Tokita Et Al 2017 Morphometric and Adaptive RadiationDocument17 pagesTokita Et Al 2017 Morphometric and Adaptive RadiationHugoBenitezdNo ratings yet
- A Synopsis of The Rubiaceae of The States of Mato Grosso and Mato Grosso Do Sul, Brazil, With A Key To Genera, and A Preliminary Species ListDocument84 pagesA Synopsis of The Rubiaceae of The States of Mato Grosso and Mato Grosso Do Sul, Brazil, With A Key To Genera, and A Preliminary Species ListJúnior SenseyNo ratings yet
- For Connect With Final Answers Gen. Int 2 in Class Lecture 6 Gene Interaction - PART 2Document33 pagesFor Connect With Final Answers Gen. Int 2 in Class Lecture 6 Gene Interaction - PART 2Anonymous Ef82K4xyNo ratings yet
- Barbara YelvertonDocument5 pagesBarbara YelvertonsendinoNo ratings yet
- Mammals in Sri LankaDocument2 pagesMammals in Sri LankaVangeesa SumanasekaraNo ratings yet
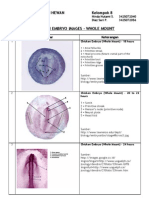
- Chick Embryo WMDocument6 pagesChick Embryo WMdhyan_ajjah67% (3)
- Cobb500 SF Breeder Management Chart Pounds PDFDocument2 pagesCobb500 SF Breeder Management Chart Pounds PDFjai10141No ratings yet
- Ministerio de Turismo: Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo ZarpoDocument10 pagesMinisterio de Turismo: Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpo Zarpocarina vrnturaNo ratings yet
- Rank The 10 Best and Most Fascinating Zoos in The WorldDocument12 pagesRank The 10 Best and Most Fascinating Zoos in The WorldTey ValenzuelaNo ratings yet
- Hazel Rhs Horticulture Level 2 Essay 1Document24 pagesHazel Rhs Horticulture Level 2 Essay 1hazeldwyerNo ratings yet
- GynoeciumDocument7 pagesGynoeciumpdekraaijNo ratings yet
- 100 Years of ZsiDocument5 pages100 Years of ZsiAshutoshkumar jhaNo ratings yet
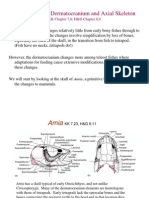
- The Dermatocranium and Axial Skeleton: KK Chapter 7,8 H&G Chapter 8,9Document25 pagesThe Dermatocranium and Axial Skeleton: KK Chapter 7,8 H&G Chapter 8,9bhavikrao7605No ratings yet
- Hygiene Que. BSS 1) What Are Insects?Document2 pagesHygiene Que. BSS 1) What Are Insects?Amaresh JhaNo ratings yet
- Epithelial TissueDocument10 pagesEpithelial Tissuememe bolongonNo ratings yet
- 2 Mora y Robinson Loxocemus - LepidochelysDocument2 pages2 Mora y Robinson Loxocemus - LepidochelysJosé MoraNo ratings yet
- Full Basic Psychopharmacology For Counselors and Psychotherapists 2Nd Edition Sinacola Test Bank PDFDocument25 pagesFull Basic Psychopharmacology For Counselors and Psychotherapists 2Nd Edition Sinacola Test Bank PDFwilliam.ely879100% (16)
- Identifying Birds by Their Beaks Lesson Plan PDFDocument3 pagesIdentifying Birds by Their Beaks Lesson Plan PDFASHISH PADHY100% (1)
- All in One Practice Sheets For ClassDocument90 pagesAll in One Practice Sheets For ClassLakshay poswalNo ratings yet
- Formicidae PDFDocument3,127 pagesFormicidae PDFCindy GarciaNo ratings yet
- 001-111 mmv76 Gomon Choerodon 7 WebDocument111 pages001-111 mmv76 Gomon Choerodon 7 WebM DayatNo ratings yet