You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- VB2014 Anatomy Etymology EbookDocument22 pagesVB2014 Anatomy Etymology EbookDado ArmawanNo ratings yet
- Neck Muscles 041715 PDFDocument15 pagesNeck Muscles 041715 PDFJohn DavidNo ratings yet
- Neck Muscles 041715 PDFDocument15 pagesNeck Muscles 041715 PDFJohn DavidNo ratings yet
- Neck Muscles 041715 PDFDocument15 pagesNeck Muscles 041715 PDFJohn DavidNo ratings yet
- Mpez-6 Installation Manual - Inline 201007Document8 pagesMpez-6 Installation Manual - Inline 201007api-244989438No ratings yet
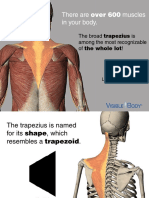
- There Are in Your Body.: Over 600 MusclesDocument14 pagesThere Are in Your Body.: Over 600 MusclesJohn DavidNo ratings yet
- Trigonometrija FormuliceDocument1 pageTrigonometrija FormuliceJohn David0% (1)
- Impact ofDocument59 pagesImpact ofJohn DavidNo ratings yet
- 1 StepenovanjeDocument7 pages1 StepenovanjeDanijela BanjevicNo ratings yet
- 6.linearne Jednacine I Nejednacine ZadaciDocument13 pages6.linearne Jednacine I Nejednacine Zadacislavkoni100% (2)
- The Human Cell 120114Document16 pagesThe Human Cell 120114jfmacNo ratings yet
- VB Muscle BURSAE 081814 PDFDocument18 pagesVB Muscle BURSAE 081814 PDFtaurito26No ratings yet
- VB Skeletal System Funfacts 2015Document21 pagesVB Skeletal System Funfacts 2015John DavidNo ratings yet
- VB BodyFactsandFiction061313 PDFDocument20 pagesVB BodyFactsandFiction061313 PDFJohn DavidNo ratings yet
- VB Build A Heart031715Document15 pagesVB Build A Heart031715John DavidNo ratings yet
- Anatomski Atlas KOSTI GLAVEDocument19 pagesAnatomski Atlas KOSTI GLAVEVesna StevanovicNo ratings yet
- VB Muscle Premium Preview 033015Document15 pagesVB Muscle Premium Preview 033015John DavidNo ratings yet
- VB AnatomyAtlas Preview 112513Document11 pagesVB AnatomyAtlas Preview 112513John DavidNo ratings yet
- Anatomy For Yogis Ebook 041816Document11 pagesAnatomy For Yogis Ebook 041816John DavidNo ratings yet
- Review Key Pelvic Structures: With Human Anatomy AtlasDocument20 pagesReview Key Pelvic Structures: With Human Anatomy AtlasAlana LoweNo ratings yet
- Sphenoid Ebook 041715Document15 pagesSphenoid Ebook 041715John DavidNo ratings yet
- AP Positions and PlanesDocument20 pagesAP Positions and PlanesBrunero Meozzi100% (1)
- Adventure Plus: Treadmill Owner'S ManualDocument28 pagesAdventure Plus: Treadmill Owner'S ManualJohn DavidNo ratings yet
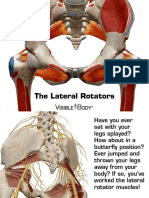
- Lateral Rotators 030714 PDFDocument12 pagesLateral Rotators 030714 PDFJohn DavidNo ratings yet
- AP Olfaction 110414 PDFDocument11 pagesAP Olfaction 110414 PDFJohn DavidNo ratings yet
- Skeleton AtlasAxis 041715Document17 pagesSkeleton AtlasAxis 041715Alaa SalamNo ratings yet
- Three Cool Facts About The: Hip BonesDocument6 pagesThree Cool Facts About The: Hip BonesAlaa SalamNo ratings yet
- A PS G: & Tudy UideDocument23 pagesA PS G: & Tudy UideBrunero MeozziNo ratings yet
- The Eye: (We'll Leave The Lord Sauron Jokes To You.)Document15 pagesThe Eye: (We'll Leave The Lord Sauron Jokes To You.)taurito26No ratings yet
- AP Prefixes 092414Document25 pagesAP Prefixes 092414Brunero MeozziNo ratings yet
- Biography of Murray (1893-1988) : PersonologyDocument6 pagesBiography of Murray (1893-1988) : PersonologyMing100% (1)
- Keandalan Bangunan Rumah SusunDocument9 pagesKeandalan Bangunan Rumah SusunDewi ARimbiNo ratings yet
- Design and Fabrication of Floor Cleaning Machine - A ReviewDocument4 pagesDesign and Fabrication of Floor Cleaning Machine - A ReviewIJIERT-International Journal of Innovations in Engineering Research and Technology100% (1)
- Factors Associated With Early Pregnancies Among Adolescent Girls Attending Selected Health Facilities in Bushenyi District, UgandaDocument12 pagesFactors Associated With Early Pregnancies Among Adolescent Girls Attending Selected Health Facilities in Bushenyi District, UgandaKIU PUBLICATION AND EXTENSIONNo ratings yet
- Rifle Threat Performance Matrix: SeriesDocument1 pageRifle Threat Performance Matrix: SeriesKuhnNo ratings yet
- Damasco - Cpi - Activity No. 10Document18 pagesDamasco - Cpi - Activity No. 10LDCU - Damasco, Erge Iris M.No ratings yet
- MLT IMLT Content Guideline 6-14Document4 pagesMLT IMLT Content Guideline 6-14Arif ShaikhNo ratings yet
- Schools Division of Roxas CityDocument4 pagesSchools Division of Roxas CityHuge Propalde EstolanoNo ratings yet
- InfertilityDocument8 pagesInfertilityrivannyNo ratings yet
- Infographic Humanistic PsychologyDocument2 pagesInfographic Humanistic Psychologyvivain.honnalli.officialNo ratings yet
- Optimization Process of Biodiesel Production With Ultrasound Assisted by Using Central Composite Design MethodsDocument47 pagesOptimization Process of Biodiesel Production With Ultrasound Assisted by Using Central Composite Design MethodsMiftahFakhriansyahNo ratings yet
- Positioning Strategy - Mineral WaterDocument2 pagesPositioning Strategy - Mineral WaterRakesh SelvamNo ratings yet
- ICGSE Chemistry Chapter 1 - The Particulate Nature of MatterDocument29 pagesICGSE Chemistry Chapter 1 - The Particulate Nature of MatterVentus TanNo ratings yet
- Prestress 3.0Document10 pagesPrestress 3.0Jonel CorbiNo ratings yet
- Health Module 6 g7Document26 pagesHealth Module 6 g7Richelle PamintuanNo ratings yet
- Cough: A Rapid Expulsion of Air FromDocument2 pagesCough: A Rapid Expulsion of Air FromaubyangNo ratings yet
- Building Technology (CE1303) : Window: Lecturer: Madam FatinDocument19 pagesBuilding Technology (CE1303) : Window: Lecturer: Madam FatinRazif AjibNo ratings yet
- ListwarehouseDocument1 pageListwarehouseKautilya KalyanNo ratings yet
- Quiz BowlDocument36 pagesQuiz BowlSherry GonzagaNo ratings yet
- Installation and Operation Manual: Proact™ Ii Electric Powered Actuator and DriverDocument32 pagesInstallation and Operation Manual: Proact™ Ii Electric Powered Actuator and DriverDjebali MouradNo ratings yet
- Iso 2281 1990Document8 pagesIso 2281 1990jesus torresNo ratings yet
- Work Environment Act (1977:1160) : Chapter 1 Purpose and Scope of The ActDocument26 pagesWork Environment Act (1977:1160) : Chapter 1 Purpose and Scope of The ActNatalie BlackmoreNo ratings yet
- NIST Standard Reference Materials® 2023 CatalogDocument128 pagesNIST Standard Reference Materials® 2023 CatalogAbdul HaseebNo ratings yet
- Tropical Fruit CHAPTER-3Document32 pagesTropical Fruit CHAPTER-3Jeylan FekiNo ratings yet
- 2-Phase Synchronous-Rectified Buck Controller For Mobile GPU PowerDocument18 pages2-Phase Synchronous-Rectified Buck Controller For Mobile GPU PowerMax Assistência TécnicaNo ratings yet
- List Acreditated JournalDocument2,124 pagesList Acreditated JournalSetiyo PrajokoNo ratings yet
- Giardiasis PDFDocument14 pagesGiardiasis PDFSaad Motawéa0% (1)
- D2C - Extensive ReportDocument54 pagesD2C - Extensive ReportVenketesh100% (1)