You might also like
- (Cambridge Studies in Philosophy and Biology) Manfred D. Laubichler, Jane Maienschein - Form and Function in Developmental Evolution. Cambridge Studies in Philosophy and Biology-Cambridge University PDocument254 pages(Cambridge Studies in Philosophy and Biology) Manfred D. Laubichler, Jane Maienschein - Form and Function in Developmental Evolution. Cambridge Studies in Philosophy and Biology-Cambridge University PAnonymous pZ2FXUycNo ratings yet
- DNA Profiling 3 (Repaired)Document59 pagesDNA Profiling 3 (Repaired)Anni FipNo ratings yet
- DNA - Recombination Interactions and Repair: Proceedings of the FEBS Symposium on DNA, Liblice, 1979From EverandDNA - Recombination Interactions and Repair: Proceedings of the FEBS Symposium on DNA, Liblice, 1979S. ZadrazilNo ratings yet
- International Workshop On "Pharmacophore Modelling and Molecular Docking Studies in Bio-Science Research"Document2 pagesInternational Workshop On "Pharmacophore Modelling and Molecular Docking Studies in Bio-Science Research"Quaxon Bio & IT SolutionsNo ratings yet
- DNA Phenotyping: - No - Ing) Is The Process of Predicting An Organism's Phenotype Using Only Genetic Information CollectedDocument5 pagesDNA Phenotyping: - No - Ing) Is The Process of Predicting An Organism's Phenotype Using Only Genetic Information CollectedNemo NemoNo ratings yet
- WoW64 Bypassing EMET PDFDocument19 pagesWoW64 Bypassing EMET PDFSatria Ady PradanaNo ratings yet
- Developmental BiologyDocument21 pagesDevelopmental BiologygahanNo ratings yet
- Clostridium Botulinum NeurotoxinsDocument12 pagesClostridium Botulinum NeurotoxinsHening Tri UtamiNo ratings yet
- Drosophila Models For Human DiseasesDocument314 pagesDrosophila Models For Human Diseasesphyaravi100% (1)
- DNA TypingDocument30 pagesDNA TypingPriyanka Pharma100% (2)
- MSC BiotechnologyDocument63 pagesMSC Biotechnologyprasanth rkNo ratings yet
- Medical Biotechnology: A Resource Guide For Biotechnology Club SponsorsDocument39 pagesMedical Biotechnology: A Resource Guide For Biotechnology Club Sponsorsim_mogerzNo ratings yet
- An Overview of Cardiac MorphogenesisDocument12 pagesAn Overview of Cardiac MorphogenesisAnonymous 9wHscMNo ratings yet
- Syllabus of Bio-Informatics, PUCCDocument14 pagesSyllabus of Bio-Informatics, PUCCMd MohiuddinNo ratings yet
- 2020 Ccaf Legal Regulatory Considerations ReportDocument50 pages2020 Ccaf Legal Regulatory Considerations ReportKarem MahmoudNo ratings yet
- Biotechnology SyllabusDocument111 pagesBiotechnology SyllabusSatyam SinghNo ratings yet
- LSM3233 Lecture 1Document43 pagesLSM3233 Lecture 1Prab NathanNo ratings yet
- An Analysis of The Intellectual Property Rights Protection Obligations and Joint Liabilities of E Commerce Platform OperatorsDocument12 pagesAn Analysis of The Intellectual Property Rights Protection Obligations and Joint Liabilities of E Commerce Platform OperatorsEditor IJTSRDNo ratings yet
- Consolidated School Quality ReportDocument108 pagesConsolidated School Quality ReportSudhir VashistNo ratings yet
- Acquisition of New GenesDocument18 pagesAcquisition of New GenesMudit Misra100% (1)
- Genetics of CancerDocument64 pagesGenetics of CancerErin HillNo ratings yet
- 6476 Formulation of Alternative Culture Media For Bacterial and Fungal GrowthDocument6 pages6476 Formulation of Alternative Culture Media For Bacterial and Fungal GrowthHanief MitsukhaNo ratings yet
- Notes On Pettit's The Consequentialist Perspective'Document3 pagesNotes On Pettit's The Consequentialist Perspective'Joseph BowmanNo ratings yet
- A Comprehensive Review of Wireless Body Area Network - 2019 - A Comprehensive Review of Wireless Body Area NetworkDocument1 pageA Comprehensive Review of Wireless Body Area Network - 2019 - A Comprehensive Review of Wireless Body Area NetworkGeneration GenerationNo ratings yet
- The Eukaryotic Genome andDocument76 pagesThe Eukaryotic Genome andKehkashan KhanNo ratings yet
- Dna Identification: K GopiDocument29 pagesDna Identification: K GopigopikupuchittyNo ratings yet
- GMS 6012 Basic Medical Genetics Syllabus Spring 2015, Blanck v4Document23 pagesGMS 6012 Basic Medical Genetics Syllabus Spring 2015, Blanck v4Ahmad AlSwaisNo ratings yet
- Forensic Science and Criminal PsychologyDocument13 pagesForensic Science and Criminal PsychologySANTOSH MANDALENo ratings yet
- This Video: To Do: Watch To Complete This TableDocument1 pageThis Video: To Do: Watch To Complete This TableLaura Cormane100% (1)
- A Geroscience Perspective On Immune Resilience and Infectious Diseases - A Potential Case For MetforminDocument20 pagesA Geroscience Perspective On Immune Resilience and Infectious Diseases - A Potential Case For MetforminAna Raquel Realista Coelho Dos Santos PedrosaNo ratings yet
- P - B I F DSP A: Olynomial Ased Nterpolation Ilters For PplicationsDocument68 pagesP - B I F DSP A: Olynomial Ased Nterpolation Ilters For PplicationsMahesh A BirariNo ratings yet
- One Day International Workshop On "Molecular Docking: From Theory To Practice"Document2 pagesOne Day International Workshop On "Molecular Docking: From Theory To Practice"Quaxon Bio & IT Solutions100% (1)
- Biotechnology and BioinformaticsDocument136 pagesBiotechnology and BioinformaticsCandy BoyNo ratings yet
- Forensic DNA TestingDocument15 pagesForensic DNA TestingMuazzam FarooqNo ratings yet
- From DNA Biosensor To Gene ChipDocument6 pagesFrom DNA Biosensor To Gene ChipramzermanNo ratings yet
- Baglivi - Practice of PhysicDocument462 pagesBaglivi - Practice of Physicglamis100% (1)
- Biosafety and Biosecurity Challenges Facing VeteriDocument11 pagesBiosafety and Biosecurity Challenges Facing VeteriJennifer Carolina Palma VanegasNo ratings yet
- Comparative Criminal Procedure - Adversial and Inuisatorial System NewDocument39 pagesComparative Criminal Procedure - Adversial and Inuisatorial System Newarchit jainNo ratings yet
- @ Current Trends in Biomedical Engineering and Bioimages AnalysisDocument344 pages@ Current Trends in Biomedical Engineering and Bioimages AnalysisNeo Peter KimNo ratings yet
- AP Psych Study GuideDocument83 pagesAP Psych Study GuideKitty chenNo ratings yet
- May 2022: Most Downloaded Articles in Computer Science &information TechnologyDocument33 pagesMay 2022: Most Downloaded Articles in Computer Science &information TechnologyAnonymous Gl4IRRjzNNo ratings yet
- Illumina Sequencing IntroductionDocument12 pagesIllumina Sequencing IntroductionSushil SinghNo ratings yet
- Single FatherDocument132 pagesSingle FatherApril Joy Andres MadriagaNo ratings yet
- Gene MutationDocument20 pagesGene Mutationdheeresh aggarwalNo ratings yet
- The Genetic CodeDocument67 pagesThe Genetic CodeRhea SharmaNo ratings yet
- Cloning Dolly & MicromanipulationDocument29 pagesCloning Dolly & Micromanipulationnitinyadav16No ratings yet
- Biotech PDFDocument34 pagesBiotech PDFSelvakapoorNo ratings yet
- Single-Nucleotide PolymorphismDocument21 pagesSingle-Nucleotide PolymorphismdearbhupiNo ratings yet
- Hales - A Primer On The Drosophila Model SystemDocument28 pagesHales - A Primer On The Drosophila Model SystemCramblerNo ratings yet
- PFAM DatabaseDocument22 pagesPFAM DatabaseNadish KumarNo ratings yet
- Evaluating Cell Fractionation Procedures and Cytochemical TestsDocument6 pagesEvaluating Cell Fractionation Procedures and Cytochemical TestsJosh Zollman100% (1)
- The Microchip World: Recent Advances and Options For Shelters and VeterinariansDocument4 pagesThe Microchip World: Recent Advances and Options For Shelters and VeterinariansJulie SavoieNo ratings yet
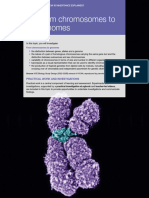
- C 06 From Chromosomes To GenomesDocument60 pagesC 06 From Chromosomes To GenomesHamadaShehataNo ratings yet
- Nano Based Approach To Combat Emerging Viral NIPAH Virus InfectionDocument26 pagesNano Based Approach To Combat Emerging Viral NIPAH Virus InfectionYolanda Charlota ThenuNo ratings yet
- Lect# Plant Molecular MarkersDocument23 pagesLect# Plant Molecular MarkersSohail Ahmed100% (1)
- PCR, RT-PCR, Nested-Pcr, Multiplex PCR, Quantitative PCRDocument116 pagesPCR, RT-PCR, Nested-Pcr, Multiplex PCR, Quantitative PCRYunizardiNo ratings yet
- Unit 2Document103 pagesUnit 2Suraj VermaNo ratings yet
- BTech Biotech FTDocument100 pagesBTech Biotech FThinderNo ratings yet
- Nexus ICSIDocument114 pagesNexus ICSIDr.Pratixa JoshiNo ratings yet
- Forensic Dna Phenotyping (FDP)Document12 pagesForensic Dna Phenotyping (FDP)ifrahkhalid52No ratings yet
- Pre TOEFL PDFDocument1 pagePre TOEFL PDFMuhammadRizalNNo ratings yet
- Muhammad Rizal Nurhadi: April 17, 2020Document1 pageMuhammad Rizal Nurhadi: April 17, 2020MuhammadRizalNNo ratings yet
- Ivig Supplement Septoct10Document12 pagesIvig Supplement Septoct10MuhammadRizalNNo ratings yet
- Rheumatoid ArthritisDocument48 pagesRheumatoid ArthritisMuhammadRizalN100% (1)
- Tabel Siklus Panklin Xxiv, XXV, Xxvi 2016Document3 pagesTabel Siklus Panklin Xxiv, XXV, Xxvi 2016MuhammadRizalNNo ratings yet
- Syphilis: Communicable Disease Management ProtocolDocument20 pagesSyphilis: Communicable Disease Management ProtocolMuhammadRizalNNo ratings yet
- Effectiveness of Manual Therapy On Chronic Pelvic PainDocument41 pagesEffectiveness of Manual Therapy On Chronic Pelvic PainMuhammadRizalN100% (1)
- Nomegestrol AcetateDocument2 pagesNomegestrol AcetateMuhammadRizalNNo ratings yet
- Treatment of Menstrual CrampsDocument3 pagesTreatment of Menstrual CrampsMuhammadRizalNNo ratings yet
- Program of ActivitiesDocument2 pagesProgram of ActivitiesVon Limuel LopezNo ratings yet
- En DAY4 David Chen Building The AI Computing Platform For Pervasive Intelligence enDocument8 pagesEn DAY4 David Chen Building The AI Computing Platform For Pervasive Intelligence endieuwrignNo ratings yet
- Lecture 5Document20 pagesLecture 5Paylaşım KanalıNo ratings yet
- Analyst - Finance, John Lewis John Lewis PartnershipDocument2 pagesAnalyst - Finance, John Lewis John Lewis Partnershipsecret_1992No ratings yet
- DLL Drafting 7Document4 pagesDLL Drafting 7Ram Dacz100% (3)
- Pamphlet 89 Chlorine Scrubbing SystemsDocument36 pagesPamphlet 89 Chlorine Scrubbing Systemshfguavita100% (4)
- (Ug, PG & PHD) Fellowship: Tih-Iot Chanakya GroupDocument3 pages(Ug, PG & PHD) Fellowship: Tih-Iot Chanakya GroupVijay M.MNo ratings yet
- Admission: North South University (NSU) Question Bank Summer 2019Document10 pagesAdmission: North South University (NSU) Question Bank Summer 2019Mahmoud Hasan100% (7)
- Hypothesis Testing Random MotorsDocument8 pagesHypothesis Testing Random MotorsLinn ArshadNo ratings yet
- GL Career Academy Data AnalyticsDocument7 pagesGL Career Academy Data AnalyticsDeveloper GuideNo ratings yet
- Moc3040 MotorolaDocument3 pagesMoc3040 MotorolaBryanTipánNo ratings yet
- DA-I Question Bank From Module 1-3 of PHY1701 Course, Winter Semester 2020-21Document6 pagesDA-I Question Bank From Module 1-3 of PHY1701 Course, Winter Semester 2020-21Likith MallipeddiNo ratings yet
- Stress: Problem SetDocument2 pagesStress: Problem SetDanielle FloridaNo ratings yet
- JCPS School Safety PlanDocument14 pagesJCPS School Safety PlanDebbie HarbsmeierNo ratings yet
- Kat-A 4102 Rotovalve Edition3!12!02-2013 enDocument4 pagesKat-A 4102 Rotovalve Edition3!12!02-2013 enWalter PiracocaNo ratings yet
- Green ThumbDocument2 pagesGreen ThumbScarlet Sofia Colmenares VargasNo ratings yet
- 448 Authors of Different Chemistry BooksDocument17 pages448 Authors of Different Chemistry BooksAhmad MNo ratings yet
- Namagunga Primary Boarding School: Primary Six Holiday Work 2021 EnglishDocument10 pagesNamagunga Primary Boarding School: Primary Six Holiday Work 2021 EnglishMonydit santinoNo ratings yet
- Use The Analysis ToolPak To Perform Complex Data Analysis - Excel - OfficeDocument5 pagesUse The Analysis ToolPak To Perform Complex Data Analysis - Excel - OfficedakingNo ratings yet
- Money Order Sauce.Document2 pagesMoney Order Sauce.randomNo ratings yet
- Corrosion Protection PT Tosanda Dwi SapurwaDocument18 pagesCorrosion Protection PT Tosanda Dwi SapurwaYoga FirmansyahNo ratings yet
- EXPERIMENT 1 - Bendo Marjorie P.Document5 pagesEXPERIMENT 1 - Bendo Marjorie P.Bendo Marjorie P.100% (1)
- Mediator Analysis of Job Embeddedness - Relationship Between Work-Life Balance Practices and Turnover IntentionsDocument15 pagesMediator Analysis of Job Embeddedness - Relationship Between Work-Life Balance Practices and Turnover IntentionsWitty MindsNo ratings yet
- Car Parking DesignDocument6 pagesCar Parking Designcharler kinyuajNo ratings yet
- Auto CadDocument67 pagesAuto CadkltowerNo ratings yet
- Per Dev Dlp-1-2 - 3 SelfDocument6 pagesPer Dev Dlp-1-2 - 3 SelfMonisa SocorinNo ratings yet
- Classroom Management PlanDocument14 pagesClassroom Management PlancowlesmathNo ratings yet
- Fiedler1950 - A Comparison of Therapeutic Relationships in PsychoanalyticDocument10 pagesFiedler1950 - A Comparison of Therapeutic Relationships in PsychoanalyticAnca-Maria CovaciNo ratings yet
- How To Install 64 Bits IDES On 32 Bits OSDocument1 pageHow To Install 64 Bits IDES On 32 Bits OSMuhammad JaveedNo ratings yet
- Changing Sentences in The Simple Present Tense Into PassiveDocument4 pagesChanging Sentences in The Simple Present Tense Into PassiveBernadette NarteNo ratings yet