You might also like
- 5 A.B. Scan InstrumentDocument13 pages5 A.B. Scan InstrumentHendrana TjahjadiNo ratings yet
- File 2Document32 pagesFile 2stanikabhiNo ratings yet
- 05 Us ArtifactsDocument33 pages05 Us ArtifactsHendrana TjahjadiNo ratings yet
- Safety AnalyzerDocument104 pagesSafety AnalyzerHendrana TjahjadiNo ratings yet
- LED NIR Spectroscope 1475 925X 4 29Document11 pagesLED NIR Spectroscope 1475 925X 4 29TheDreamMNo ratings yet
- Bci Whellchair 2015Document6 pagesBci Whellchair 2015Hendrana TjahjadiNo ratings yet
- Embedded System in Biomedical Applications: Challenges AheadDocument3 pagesEmbedded System in Biomedical Applications: Challenges AheadHendrana TjahjadiNo ratings yet
- File 2Document32 pagesFile 2stanikabhiNo ratings yet
- Microprocessor-Based System: Ambulatory Ecg MonitoringDocument2 pagesMicroprocessor-Based System: Ambulatory Ecg MonitoringHendrana TjahjadiNo ratings yet
- ETHZDocument28 pagesETHZIepurash MadalinaNo ratings yet
- BloodPressure AliasingDocument35 pagesBloodPressure AliasingHendrana TjahjadiNo ratings yet
- A Real-Time Microprocessor QRS DetectorDocument9 pagesA Real-Time Microprocessor QRS DetectorHendrana TjahjadiNo ratings yet
- Ceiling Pendant: WWW - Mmakorea.co - KRDocument8 pagesCeiling Pendant: WWW - Mmakorea.co - KRHendrana TjahjadiNo ratings yet
- A Real-Time Microprocessor QRS DetectorDocument9 pagesA Real-Time Microprocessor QRS DetectorHendrana TjahjadiNo ratings yet
- Jurnal Sensor Spo2.4Document131 pagesJurnal Sensor Spo2.4Hendrana TjahjadiNo ratings yet
- Jurnal Firmware Spo2.5Document8 pagesJurnal Firmware Spo2.5Hendrana Tjahjadi0% (1)
- Jurnal Hardware Spo2.2Document12 pagesJurnal Hardware Spo2.2Hendrana TjahjadiNo ratings yet
- Bali GpsDocument38 pagesBali GpsHendrana TjahjadiNo ratings yet
- EndosDocument50 pagesEndosBMT100% (3)
- PPGDocument7 pagesPPGHendrana TjahjadiNo ratings yet
- Pulse Oximeter Principle 1Document8 pagesPulse Oximeter Principle 1Ell ElyanaNo ratings yet
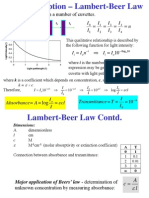
- Beer Lambert Oximeter.2Document5 pagesBeer Lambert Oximeter.2Hendrana TjahjadiNo ratings yet
- Part 3 Using Light For DetectionDocument48 pagesPart 3 Using Light For DetectionJosé ClementeNo ratings yet
- Berlin Dve - en PDFDocument2 pagesBerlin Dve - en PDFHendrana TjahjadiNo ratings yet
- Maintenance Form: Anesthesia UnitDocument2 pagesMaintenance Form: Anesthesia UnitHendrana TjahjadiNo ratings yet
- Bisnis Plan Spo2Document37 pagesBisnis Plan Spo2Hendrana TjahjadiNo ratings yet
- Price List AlfakesDocument5 pagesPrice List AlfakesHendrana TjahjadiNo ratings yet
- Inspection Form: Dept/Loc: Merk/Type: S/N: No. Inventory: InspectorDocument2 pagesInspection Form: Dept/Loc: Merk/Type: S/N: No. Inventory: InspectorHendrana TjahjadiNo ratings yet
- Understanding Why Hospitals Outsource Biomed Services: Observations and InsightsDocument3 pagesUnderstanding Why Hospitals Outsource Biomed Services: Observations and InsightsHendrana TjahjadiNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- History of NeuroscienceDocument17 pagesHistory of NeurosciencerazorviperNo ratings yet
- Tigers SensesDocument5 pagesTigers SensespaidalaNo ratings yet
- BrainstemA AtlasDocument23 pagesBrainstemA AtlasAndrada CatrinoiuNo ratings yet
- Control of Pain in Operative DentistryDocument5 pagesControl of Pain in Operative DentistryAnkit VirdiNo ratings yet
- A Review of Neuromuscular MonitoringDocument10 pagesA Review of Neuromuscular Monitoringdrcds99No ratings yet
- EMG Lab Simple Muscle TwitchDocument8 pagesEMG Lab Simple Muscle TwitchShauki AliNo ratings yet
- Science Olympiad: Anatanmy 2013 - Div BDocument45 pagesScience Olympiad: Anatanmy 2013 - Div BSuper_Star99No ratings yet
- Sensory PhysiologyDocument57 pagesSensory PhysiologyeryxspNo ratings yet
- Physiological Basis of BehaviorDocument35 pagesPhysiological Basis of BehaviorAnna Rhea BurerosNo ratings yet
- Taste and SmellDocument36 pagesTaste and SmellKarthikeyan Balakrishnan100% (2)
- Where Is The LesionDocument12 pagesWhere Is The LesionHo Yong WaiNo ratings yet
- Music, Cognition, and Computerized Sound, Cook, P.R., 1999Document379 pagesMusic, Cognition, and Computerized Sound, Cook, P.R., 1999George Roldugin100% (3)
- The Student's Guide To Cognitive Neuroscience: Second EditionDocument3 pagesThe Student's Guide To Cognitive Neuroscience: Second EditionslowlightNo ratings yet
- Outline Learning Objectives and Contingency Plan Nervous and SensoryDocument3 pagesOutline Learning Objectives and Contingency Plan Nervous and Sensoryapi-301903881No ratings yet
- 4 - Therapeutic Effects of Low Frequency Currents On TissuesIVDocument52 pages4 - Therapeutic Effects of Low Frequency Currents On TissuesIVhuxley2378No ratings yet
- Theories of Colour VisionDocument4 pagesTheories of Colour VisionZumairi Tamin0% (1)
- Unlocking The BrainDocument3 pagesUnlocking The BrainPsychedelicBaronNo ratings yet
- Unit5 Prac QsDocument43 pagesUnit5 Prac QsLola ShmeerNo ratings yet
- Functions of The Nervous SystemDocument5 pagesFunctions of The Nervous SystemJungkookie BaeNo ratings yet
- How Exercise Can Calm AnxietyDocument3 pagesHow Exercise Can Calm AnxietyAlejandro FernadnezNo ratings yet
- SynapsesDocument46 pagesSynapseshan abuur ahmedNo ratings yet
- BIO 3303 Final Exam ReviewDocument31 pagesBIO 3303 Final Exam ReviewSara YuenNo ratings yet
- Hebbian LearningDocument53 pagesHebbian LearningPiyush BansalNo ratings yet
- The Sensorimotor System Part I The Physiologic Basis of Functional Joint StablityDocument9 pagesThe Sensorimotor System Part I The Physiologic Basis of Functional Joint StablityMauro Esban Obreque QuianNo ratings yet
- 1-5 Hear N SightDocument19 pages1-5 Hear N SightNor ShuhadaNo ratings yet
- A Chimpanzee BrainDocument21 pagesA Chimpanzee BrainKevin ChenNo ratings yet
- Cap II - Curs 5 - 6 - Somatosensory System - 2014 - 2015 - PDFDocument137 pagesCap II - Curs 5 - 6 - Somatosensory System - 2014 - 2015 - PDFMaria MadalinaNo ratings yet
- Limbic Sys - ArchDocument85 pagesLimbic Sys - Archdrkadiyala2No ratings yet
- Mecanismos Celulares de La Neuroplasticidad Sin ProteccionDocument22 pagesMecanismos Celulares de La Neuroplasticidad Sin ProteccionMati Sanabria100% (1)