You might also like
- MTHFR 97d077a0a2e9 20200116170647 PDFDocument31 pagesMTHFR 97d077a0a2e9 20200116170647 PDFOpen PrizesNo ratings yet
- Food & Genes: Term Paper By: Phurpa D. ThungonDocument15 pagesFood & Genes: Term Paper By: Phurpa D. ThungonMehul BhardwajNo ratings yet
- DR - Amy Yasko Microbes Metals MethylationDocument211 pagesDR - Amy Yasko Microbes Metals Methylationmaimutalin100% (2)
- Yasko-General Important Information To Guide You On Your Road To WellnessDocument31 pagesYasko-General Important Information To Guide You On Your Road To WellnessMichael CabarlesNo ratings yet
- Cell Nutrition GenesDocument24 pagesCell Nutrition GenesAlberto Alberto100% (3)
- Nature Outlook Nutrigenomics. How Food Shapes Health and Evolution - TextmarkDocument43 pagesNature Outlook Nutrigenomics. How Food Shapes Health and Evolution - Textmarktamy_001No ratings yet
- Methylation DiagramDocument1 pageMethylation Diagrampkdo2003No ratings yet
- ION Guide 2006Document16 pagesION Guide 2006Metametrix100% (2)
- Endocrine HarmonyDocument68 pagesEndocrine HarmonyafnanNo ratings yet
- Dietary Supplement Fact SheetDocument25 pagesDietary Supplement Fact SheetAngga SebagustionnesNo ratings yet
- Fmu Trainging ModuleDocument23 pagesFmu Trainging ModulesamNo ratings yet
- How to Balance Your Homocysteine the Right Way - The Genetic Advantage: The genetic advantage, #2From EverandHow to Balance Your Homocysteine the Right Way - The Genetic Advantage: The genetic advantage, #2No ratings yet
- MTHFR GeneDocument3 pagesMTHFR GeneMili LCNo ratings yet
- Threebooksoccult PDFDocument623 pagesThreebooksoccult PDFPanphilaNo ratings yet
- Kirk Man Beginners Guide WebDocument40 pagesKirk Man Beginners Guide WebSophia Airall-ThomasNo ratings yet
- Digestion and Absorption of LipidsDocument5 pagesDigestion and Absorption of LipidsAngelo Jude Cobacha100% (1)
- PROTEINSDocument17 pagesPROTEINSAbhijat Jha100% (1)
- Biology OxidationDocument50 pagesBiology Oxidationderhangker100% (3)
- Glycogenolysis Basic PDFDocument25 pagesGlycogenolysis Basic PDFSamuel Morales Navarro100% (1)
- The Significance of The Metabolism of The Neurohormone MelatoninDocument11 pagesThe Significance of The Metabolism of The Neurohormone MelatoninDaniel Díaz CarrascoNo ratings yet
- Lipid Absorption and Transport UptakeDocument218 pagesLipid Absorption and Transport UptakeSimra Zahid100% (1)
- Somatotopic Representation of Action Words in Human Motor and Premotor CortexDocument7 pagesSomatotopic Representation of Action Words in Human Motor and Premotor CortexDr. Amy YaskoNo ratings yet
- B1 ThiamineDocument8 pagesB1 ThiamineK AnjaliNo ratings yet
- Microbiology Sheet 8Document14 pagesMicrobiology Sheet 8Nour MohammedNo ratings yet
- Modulation of Synaptic Function by VAC14, A Protein That Regulates The Phosphoinositides PI (3,5) P2 and PI (5) PDocument15 pagesModulation of Synaptic Function by VAC14, A Protein That Regulates The Phosphoinositides PI (3,5) P2 and PI (5) PDr. Amy YaskoNo ratings yet
- Biodiversity and Functional Genomics in The Human Microbiome PDFDocument8 pagesBiodiversity and Functional Genomics in The Human Microbiome PDFabcder1234No ratings yet
- What Is DetoxificationDocument6 pagesWhat Is DetoxificationLaila AbdulNo ratings yet
- Endocrine DisruptorsDocument50 pagesEndocrine DisruptorsSnowangeleyes AngelNo ratings yet
- Methylation Part 2Document266 pagesMethylation Part 2MelissaNo ratings yet
- MTHFR Support Australia Sulphur Containing FoodsDocument2 pagesMTHFR Support Australia Sulphur Containing FoodsLisha HarbNo ratings yet
- Masterjohn - Vit D Toxicity Redefined - Vit K Molecular MechanismDocument9 pagesMasterjohn - Vit D Toxicity Redefined - Vit K Molecular MechanismBradNo ratings yet
- Start Low and Go Slow 1-21-2014Document5 pagesStart Low and Go Slow 1-21-2014Ed JonesNo ratings yet
- 13 Antacids and Controllers UpdDocument63 pages13 Antacids and Controllers Updone_nd_onlyuNo ratings yet
- Nutrigenomics The Genome Food InterfaceDocument8 pagesNutrigenomics The Genome Food InterfaceElisua G.I.No ratings yet
- Oxidative Stress and AntioxidantsDocument34 pagesOxidative Stress and AntioxidantsTauseefNo ratings yet
- MTHFR MutationDocument1 pageMTHFR MutationRobin KallisNo ratings yet
- Final Calcium Magnesium Ratio 3 1 12 PDFDocument2 pagesFinal Calcium Magnesium Ratio 3 1 12 PDFHitesh Verma100% (1)
- Yasko Methylation Pathway: Methionine Cycle Folate CycleDocument6 pagesYasko Methylation Pathway: Methionine Cycle Folate CycleEric Ehle100% (1)
- Chap 7 Organic Chem WorksheetDocument6 pagesChap 7 Organic Chem WorksheetULFA TUFFAHATINo ratings yet
- Steroidal Hormones (Testosterone, Progesterone, Estrogen PDFDocument23 pagesSteroidal Hormones (Testosterone, Progesterone, Estrogen PDFJazab ChohanNo ratings yet
- Assignment Of: Group VDocument13 pagesAssignment Of: Group VArslan Ejaz100% (1)
- Integration of Metabolism 2019Document68 pagesIntegration of Metabolism 2019Cahyani Tiara Safitri100% (1)
- Liver DetoxificationDocument22 pagesLiver DetoxificationJon SandersonNo ratings yet
- Metalloenzymes: From Bench to BedsideFrom EverandMetalloenzymes: From Bench to BedsideClaudiu T. SupuranNo ratings yet
- 8 Coenzymes and Vitamins (كيمياء حيوية صيدلانية (1 PDFDocument74 pages8 Coenzymes and Vitamins (كيمياء حيوية صيدلانية (1 PDFmaher100% (1)
- Allen's Key Note of Homeopathic Materia MedicaDocument177 pagesAllen's Key Note of Homeopathic Materia MedicaDinesh Dhoke100% (3)
- Role of Magnesium and CalciumDocument5 pagesRole of Magnesium and CalciumsemiNo ratings yet
- Rubidium-Induced Increase in Shock-Elicited Aggression in RatsDocument11 pagesRubidium-Induced Increase in Shock-Elicited Aggression in RatsDr. Amy YaskoNo ratings yet
- 12~19 생화학 테뱅 정리Document117 pages12~19 생화학 테뱅 정리조주상No ratings yet
- Mineral Metabolism and Abnormalities: Le Duong Hoang Huy M.D Email: Huyldh@pnt - Edu.vnDocument66 pagesMineral Metabolism and Abnormalities: Le Duong Hoang Huy M.D Email: Huyldh@pnt - Edu.vnLam NgoNo ratings yet
- Biomolecules (Carbohydrates, Proteins, Lipids and Nucleic Acid)Document95 pagesBiomolecules (Carbohydrates, Proteins, Lipids and Nucleic Acid)Glaiza GlaizaNo ratings yet
- Oxidative Damage, Aging and Anti-Aging Strategies PDFDocument17 pagesOxidative Damage, Aging and Anti-Aging Strategies PDFsarahNo ratings yet
- Genetic Genie Methylation ProfileDocument6 pagesGenetic Genie Methylation ProfileadamNo ratings yet
- Principles of Fish NutritionDocument21 pagesPrinciples of Fish NutritionAndi Keyen50% (2)
- The Key-Note System - Henry N. Guernsey, MDDocument7 pagesThe Key-Note System - Henry N. Guernsey, MDPanphilaNo ratings yet
- Non-Essential Amino Acids Nitrogen MetabolismDocument21 pagesNon-Essential Amino Acids Nitrogen Metabolismpradeep36No ratings yet
- Methylenetetrahydrofolate Reductase (MTHFR) Mutation DetectionDocument2 pagesMethylenetetrahydrofolate Reductase (MTHFR) Mutation DetectionRakeshKumarNo ratings yet
- DR AmyYaskoAndRichVanKProtocolDetoxDocument6 pagesDR AmyYaskoAndRichVanKProtocolDetoxCheryl Benson67% (3)
- Nutrigenomics DefinitionDocument19 pagesNutrigenomics DefinitionLutfi Aulia SupriyadiNo ratings yet
- Matabolic PathwaysDocument11 pagesMatabolic PathwaysLevi100% (2)
- Brain InvadersDocument2 pagesBrain InvadersMark HayesNo ratings yet
- Cellular Respiration: MitochondriaDocument8 pagesCellular Respiration: MitochondriaMaryRoseTrajada100% (1)
- Thorn Eh and OutDocument24 pagesThorn Eh and OutNick MaxNo ratings yet
- PC - DR - Nutrient Core - Report - (CP00053651) PDFDocument22 pagesPC - DR - Nutrient Core - Report - (CP00053651) PDFAmanda GrantNo ratings yet
- 2nd Peroxisome Metabolism 20760414Document56 pages2nd Peroxisome Metabolism 20760414Rawbeena RamtelNo ratings yet
- Nutraceutical ExecutiveSummary PDFDocument13 pagesNutraceutical ExecutiveSummary PDFAnonymous AZCGD8Uv2No ratings yet
- Margaret Lucy Tyler - How Not To Do ItDocument11 pagesMargaret Lucy Tyler - How Not To Do ItPanphilaNo ratings yet
- HYDROCARBONSDocument17 pagesHYDROCARBONSsarahNo ratings yet
- Nutrients of Bitter GourdDocument2 pagesNutrients of Bitter GourdEva Arian JoyNo ratings yet
- LH Fact Sheet Chemical Resistance 03-2019Document4 pagesLH Fact Sheet Chemical Resistance 03-2019Boro TerakolNo ratings yet
- Constitution of Sovereignty: The Summary: Submitted By: Kervy Jay T. AgraviadorDocument7 pagesConstitution of Sovereignty: The Summary: Submitted By: Kervy Jay T. AgraviadorKervy Jay AgraviadorNo ratings yet
- Section XIV - Lipid TransportDocument2 pagesSection XIV - Lipid TransportKatharine NervaNo ratings yet
- Organic Molecule:: Nucleic AcidsDocument26 pagesOrganic Molecule:: Nucleic AcidsCharles SalcedoNo ratings yet
- Studi Kualitatif Dan Kuantitatif Fitokimia Ekstrak Air Dan Ekstrak Etanol Daun Kirinyuh (Chromolaena Odorata L.) Yang Tumbuh Di Provinsi BaliDocument5 pagesStudi Kualitatif Dan Kuantitatif Fitokimia Ekstrak Air Dan Ekstrak Etanol Daun Kirinyuh (Chromolaena Odorata L.) Yang Tumbuh Di Provinsi BaliOkta OliviaNo ratings yet
- Biomolecules: Carbohydrates, Proteins, Lipids, Nucleic Acids, EnzymesDocument11 pagesBiomolecules: Carbohydrates, Proteins, Lipids, Nucleic Acids, EnzymesJemiahNo ratings yet
- Pygas Upgrading For European Steam Crackers: Joseph C Gentry and Meijuan ZengDocument5 pagesPygas Upgrading For European Steam Crackers: Joseph C Gentry and Meijuan ZengArash AbbasiNo ratings yet
- So TabDocument22 pagesSo TabdewakusumaNo ratings yet
- Class 11 Organic Basics + IUPAC Notes PDFDocument37 pagesClass 11 Organic Basics + IUPAC Notes PDFproud vedan laxmiNo ratings yet
- Comprehensive Review of Important Analytical Reagents Used in SpectrophotometryDocument29 pagesComprehensive Review of Important Analytical Reagents Used in SpectrophotometryvarishNo ratings yet
- The Solution in Energy CuringDocument22 pagesThe Solution in Energy CuringDr. Suresh ShisodiaNo ratings yet
- MBM Home Quiz BEFORE MIDTERMDocument38 pagesMBM Home Quiz BEFORE MIDTERMbhagavan prasadNo ratings yet
- Lap Mut As I Stock GudangDocument41 pagesLap Mut As I Stock Gudangputri rezeki br manurungNo ratings yet
- Zat Kimia Pembantu (Slide Kuliah II)Document55 pagesZat Kimia Pembantu (Slide Kuliah II)Abdul Rohman Heryadi100% (1)
- Vitamin A RM COADocument2 pagesVitamin A RM COAASHOK KUMAR LENKANo ratings yet
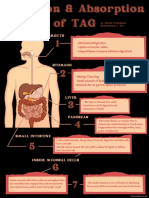
- Digestion & Absorption of TAGDocument1 pageDigestion & Absorption of TAGJanine Franchesca SuministradoNo ratings yet
- Biology InvestigatoryDocument12 pagesBiology Investigatoryalan ChackoNo ratings yet
- Methods of Synthesis of Deuterium-Labelled LipidsDocument13 pagesMethods of Synthesis of Deuterium-Labelled LipidsDavid SweedlerNo ratings yet
- Organic Chemistry Revision Questions-1Document3 pagesOrganic Chemistry Revision Questions-1jatin2006gamil.comNo ratings yet
- Repacking Licence For Web March2020 3Document11 pagesRepacking Licence For Web March2020 3aiktiplarNo ratings yet
- Industrial Organic ChemistryDocument30 pagesIndustrial Organic ChemistryChaiwatTippuwanan0% (1)
- Chemistry of Oleochemicals and SurfactantsDocument35 pagesChemistry of Oleochemicals and SurfactantsUtkarsh MankarNo ratings yet
- Aaj Ki Chemistry Ki Class..Document9 pagesAaj Ki Chemistry Ki Class..prashant panwarNo ratings yet