You might also like
- Primer Parcial IBMC-2015Document4 pagesPrimer Parcial IBMC-2015LaraBen79100% (1)
- Presentacion Geralk Naturaleza Del GenDocument28 pagesPresentacion Geralk Naturaleza Del GenWaldo Guillermo RuizNo ratings yet
- Pauta Solemne 1 BIOL240 2nd SemestreDocument9 pagesPauta Solemne 1 BIOL240 2nd SemestreLeonidas LarrondoNo ratings yet
- Solemnes Biología MolecularDocument12 pagesSolemnes Biología MolecularLeonidas LarrondoNo ratings yet
- Laboratorio 3 - Mutagenesis WannerDocument12 pagesLaboratorio 3 - Mutagenesis Wannerjorge fravega ammannNo ratings yet
- Probtransregulacion y Su Rtas.Document20 pagesProbtransregulacion y Su Rtas.Beatriz SaidmanNo ratings yet
- Problemario DNA y RNADocument3 pagesProblemario DNA y RNAJoel GomezNo ratings yet
- Preguntas Biología MolecularDocument13 pagesPreguntas Biología MolecularjaNo ratings yet
- Solemne 3 BIOL240 2021Document7 pagesSolemne 3 BIOL240 2021Leonidas LarrondoNo ratings yet
- 2 GUÍA B 2022 RespDocument31 pages2 GUÍA B 2022 RespAdrhyss FlorssNo ratings yet
- ProblemasDocument2 pagesProblemasLaura Castaño Aguirre100% (1)
- Pautas Solemne 1 y 2 Biol240Document10 pagesPautas Solemne 1 y 2 Biol240Leonidas LarrondoNo ratings yet
- TALLER - MutacionesDocument3 pagesTALLER - MutacionesJhonnathan RodriguezNo ratings yet
- I Taller Evaluativo BioquímicaDocument7 pagesI Taller Evaluativo BioquímicaCristian ArizaNo ratings yet
- Examen Entrada MEDICINADocument2 pagesExamen Entrada MEDICINAMaria Fernanda Delgado CisnerosNo ratings yet
- Solemne III Biologia Molecular 2020Document13 pagesSolemne III Biologia Molecular 2020romulo remoNo ratings yet
- TALLER 3 - Replicación - ReparaciónDocument3 pagesTALLER 3 - Replicación - ReparaciónIrene BecerraNo ratings yet
- Genetica Seminario 2Document5 pagesGenetica Seminario 2Lucia RamirezNo ratings yet
- Taller 9. Diseño de Primers 2021Document14 pagesTaller 9. Diseño de Primers 2021CarlosNo ratings yet
- Ejercicio Práctico ClonaciónDocument4 pagesEjercicio Práctico Clonaciónjhonatan ascanioNo ratings yet
- Parcial BCM Turno 2 C 09 - 23Document9 pagesParcial BCM Turno 2 C 09 - 23victor arrietaNo ratings yet
- Informe PCRDocument5 pagesInforme PCRDaniel OrtizNo ratings yet
- PCR FundamentoDocument10 pagesPCR FundamentoEillenSáezNo ratings yet
- Taller 4 - 2410Document4 pagesTaller 4 - 2410Paula Sophía CastiblancoNo ratings yet
- Guía de Problemas - 1ra Parte - BCM Nutricion 2023 - Clase 1Document8 pagesGuía de Problemas - 1ra Parte - BCM Nutricion 2023 - Clase 1leonardo farioliNo ratings yet
- Guia de Clase 4. TRADUCCIONDocument4 pagesGuia de Clase 4. TRADUCCIONMaria PalaciosNo ratings yet
- Examen Diciembre 2015Document15 pagesExamen Diciembre 2015notcaia7No ratings yet
- Practica 5 IIDocument6 pagesPractica 5 IITifanny Cuellar RamosNo ratings yet
- Biología MolecularDocument12 pagesBiología MolecularAna Laura HernándezNo ratings yet
- Cuestiones de Aplicación PCRDocument8 pagesCuestiones de Aplicación PCRLara Vázquez FafiánNo ratings yet
- Biologia Molecular Informe 2Document7 pagesBiologia Molecular Informe 2LAURA DANIELA PIÑEROS PIÑEROSNo ratings yet
- Seminario Biologia MolecularDocument9 pagesSeminario Biologia MolecularIuliaNo ratings yet
- Parcial Genética 2020 C - RDocument14 pagesParcial Genética 2020 C - RGuille DiazNo ratings yet
- Informe Caso 2 - DiagnósticoDocument5 pagesInforme Caso 2 - Diagnósticovictor arrietaNo ratings yet
- Lección 1Document7 pagesLección 1Vanesa GacioNo ratings yet
- Taller TraduccionDocument3 pagesTaller Traduccionnatalia andreaNo ratings yet
- La Fosforilación Del Núcleo de La Histona Regula La Accesibilidad Del AdnDocument5 pagesLa Fosforilación Del Núcleo de La Histona Regula La Accesibilidad Del AdneverNo ratings yet
- Examen Bioquimica Parcial3 URJC MedicinaDocument7 pagesExamen Bioquimica Parcial3 URJC MedicinaLara Fernández PedreiraNo ratings yet
- 1er Parcial IBMC 2015 - Respuestas EsperadasDocument3 pages1er Parcial IBMC 2015 - Respuestas EsperadasLaraBen79No ratings yet
- Examen 20 de Diciembre - PROTOTIPO B - MARCADO 3Document14 pagesExamen 20 de Diciembre - PROTOTIPO B - MARCADO 3estudosdamari13No ratings yet
- 888Document10 pages888api-627823169No ratings yet
- Examenes Genética I 2016 PDFDocument29 pagesExamenes Genética I 2016 PDFRicardo MunozNo ratings yet
- Guia de Ejercicios IGMODocument8 pagesGuia de Ejercicios IGMOSusana Cárdenas JiménezNo ratings yet
- TALLER 6 - Transcripción IDocument6 pagesTALLER 6 - Transcripción ILina HernandezNo ratings yet
- Taller de Recuperacion BiologíaDocument6 pagesTaller de Recuperacion BiologíaJuanEstebanMoraNo ratings yet
- BiologiaDocument8 pagesBiologiapuchiti1234No ratings yet
- Introducción de La Técnica PCRDocument6 pagesIntroducción de La Técnica PCRAngie CeronNo ratings yet
- Practica 3 Amplificacion Por PCRDocument3 pagesPractica 3 Amplificacion Por PCRIudalee Miriamm K A100% (1)
- Pau 7reptranstradDocument3 pagesPau 7reptranstradHala DeyaeNo ratings yet
- Unidad 2 - Paso 5 - Resolución de EjerciciosDocument6 pagesUnidad 2 - Paso 5 - Resolución de EjerciciosKaren Ruiz “Danna”100% (1)
- Examen BCM 21 de Febrero PROTOTIPO BDocument17 pagesExamen BCM 21 de Febrero PROTOTIPO B24laramacaNo ratings yet
- Ejercicios de GenéticaDocument32 pagesEjercicios de GenéticaSheila GarcíaNo ratings yet
- TAREA 1. Introducción GenómicaDocument8 pagesTAREA 1. Introducción GenómicaMariana MezaNo ratings yet
- Junio 2021Document9 pagesJunio 2021Sebas MamaNo ratings yet
- Recu GeneticaDocument6 pagesRecu Genetica24022003feaNo ratings yet
- Genetic A Bacterian ADocument92 pagesGenetic A Bacterian APuri JiménezNo ratings yet
- Cuestionario Cuestionario Acidos NucleicosDocument3 pagesCuestionario Cuestionario Acidos NucleicosNandoFernandoNo ratings yet
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSFrom EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSRating: 5 out of 5 stars5/5 (1)
- Atlas de ciliados y otros microorganismos frecuentes en plantas de sistemas aerobio de aguas residualesFrom EverandAtlas de ciliados y otros microorganismos frecuentes en plantas de sistemas aerobio de aguas residualesNo ratings yet
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017From EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017No ratings yet
- Refrigeracion y Congelacion 1Document18 pagesRefrigeracion y Congelacion 1Jhamer LSNo ratings yet
- II-17 Guía 08 2da Parte UnificadaDocument18 pagesII-17 Guía 08 2da Parte UnificadaHildorienNo ratings yet
- Pie Equino VaroDocument27 pagesPie Equino VaroPablo AguilarNo ratings yet
- Glosario SSDocument3 pagesGlosario SSemilioNo ratings yet
- La Investigación de La Hemoglobina y Los Orígenes de La Medicina MolecularDocument27 pagesLa Investigación de La Hemoglobina y Los Orígenes de La Medicina MolecularsalvadorNo ratings yet
- Resumen AnalgesicoDocument8 pagesResumen AnalgesicoRamarga GarciaNo ratings yet
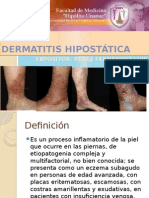
- Dermatitis HipostáticaDocument14 pagesDermatitis HipostáticaLuis Angel Perez FernandezNo ratings yet
- CapituloDocument53 pagesCapituloAnelizNo ratings yet
- Bioprocesos AnimalesDocument6 pagesBioprocesos Animalesgiovanni2013No ratings yet
- Procesos Bioquímicos de Los AlimentosDocument10 pagesProcesos Bioquímicos de Los AlimentosElio Carreño100% (3)
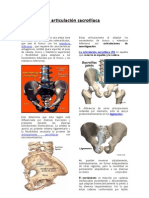
- Ajuste de Articulacion Sacroiliac ADocument7 pagesAjuste de Articulacion Sacroiliac Agustavogomez4505No ratings yet
- La Leche de VacaDocument34 pagesLa Leche de VacaJuanNo ratings yet
- Sistema Nervioso Periférico SomáticoDocument3 pagesSistema Nervioso Periférico SomáticoYerlypeña MottaNo ratings yet
- Manual Naturista Del Cancer Terapias de Qi Gong Maestro Wong Kiew KitDocument44 pagesManual Naturista Del Cancer Terapias de Qi Gong Maestro Wong Kiew KitJeffrey JenningsNo ratings yet
- Embriologia Dental ResumenDocument6 pagesEmbriologia Dental ResumenLuis FerNo ratings yet
- Citometría HemáticaDocument44 pagesCitometría HemáticaQFB 8010No ratings yet
- Tema 3 - Bases Fisiológicas Del Aprendizaje.Document37 pagesTema 3 - Bases Fisiológicas Del Aprendizaje.Mari Carmen80% (45)
- Taxonomia ParasitosDocument8 pagesTaxonomia ParasitosLuis Tovilla89% (9)
- Biología Curtis Capítulo 4Document4 pagesBiología Curtis Capítulo 4Felipe rodriguez islas0% (1)
- Extensiones SanguíneasDocument2 pagesExtensiones SanguíneasBorja GarcíaNo ratings yet
- 10 Tipos de DrogasDocument4 pages10 Tipos de DrogasLopez DanyNo ratings yet
- Ejercicios para CuadricepsDocument8 pagesEjercicios para CuadricepsJuan FranciscoNo ratings yet
- La Arcilla Patagónica - Usos MedicinalesDocument5 pagesLa Arcilla Patagónica - Usos MedicinalescarodiaNo ratings yet
- El CloroplastoDocument2 pagesEl CloroplastoChristian Fernando Rodriguez JimenesNo ratings yet
- 17 Paleopatoogia OralDocument11 pages17 Paleopatoogia OralhumbertoNo ratings yet
- Guía Aparato DigestivoDocument3 pagesGuía Aparato DigestivoEdith GuevaraNo ratings yet
- DesfibriladorDocument18 pagesDesfibriladorJoseph WarNo ratings yet
- PoriferosDocument4 pagesPoriferosAndres Felipe BermudezNo ratings yet
- Seppo AroDocument2 pagesSeppo AroAnaCastillo75% (4)
- ScaDocument98 pagesScaJavier Martinez CuellarNo ratings yet