You might also like
- Canalizaciones Discos SolaresDocument67 pagesCanalizaciones Discos Solaresperla_belmonteNo ratings yet
- El Continente AmericanoDocument2 pagesEl Continente AmericanoPavel Martinez0% (1)
- Exploracion de SueloDocument21 pagesExploracion de Sueloenmanuel tisnado uminaNo ratings yet
- Material para Matemática - Cartillas de Números Del 11 Al 20Document2 pagesMaterial para Matemática - Cartillas de Números Del 11 Al 20TaniaZayuriRosalesOrtiz100% (1)
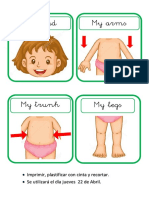
- My Body Girl (Jueves 22)Document1 pageMy Body Girl (Jueves 22)TaniaZayuriRosalesOrtizNo ratings yet
- My Body - Boy (Jueves 22)Document1 pageMy Body - Boy (Jueves 22)TaniaZayuriRosalesOrtizNo ratings yet
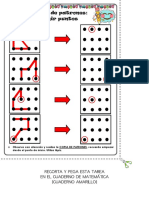
- Tarea Discriminación Visual - Copia de Patrones (Unir Puntos) (Matemática-Cuaderno Amarillo) 2Document1 pageTarea Discriminación Visual - Copia de Patrones (Unir Puntos) (Matemática-Cuaderno Amarillo) 2TaniaZayuriRosalesOrtizNo ratings yet
- Tarea Consonante P - Conocemos y Delineamos (Comunicación-Cuaderno Celeste)Document1 pageTarea Consonante P - Conocemos y Delineamos (Comunicación-Cuaderno Celeste)TaniaZayuriRosalesOrtizNo ratings yet
- Comunicado Inicio de Talleres - Jardincito de Miss Heidi (4 Años)Document2 pagesComunicado Inicio de Talleres - Jardincito de Miss Heidi (4 Años)TaniaZayuriRosalesOrtizNo ratings yet
- Materiales para Matemática - Asociar A La Cantidad (11 Al 20)Document4 pagesMateriales para Matemática - Asociar A La Cantidad (11 Al 20)TaniaZayuriRosalesOrtizNo ratings yet
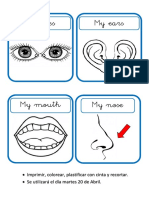
- Little Face (Martes 20)Document1 pageLittle Face (Martes 20)TaniaZayuriRosalesOrtizNo ratings yet
- Tarea Discriminación Visual - Copia de Patrones (Unir Puntos) (Matemática-Cuaderno Amarillo) 2Document1 pageTarea Discriminación Visual - Copia de Patrones (Unir Puntos) (Matemática-Cuaderno Amarillo) 2TaniaZayuriRosalesOrtizNo ratings yet
- Material para Matemática - Cartillas de Números Del 11 Al 20Document2 pagesMaterial para Matemática - Cartillas de Números Del 11 Al 20TaniaZayuriRosalesOrtizNo ratings yet
- Materiales para Matemática - Asociar A La Cantidad (11 Al 20)Document4 pagesMateriales para Matemática - Asociar A La Cantidad (11 Al 20)TaniaZayuriRosalesOrtizNo ratings yet
- Tarea Consonante P - Conocemos y Delineamos (Comunicación-Cuaderno Celeste)Document1 pageTarea Consonante P - Conocemos y Delineamos (Comunicación-Cuaderno Celeste)TaniaZayuriRosalesOrtizNo ratings yet
- Material para Matemática-Secuencia Por Color y Por FormaDocument6 pagesMaterial para Matemática-Secuencia Por Color y Por FormaTaniaZayuriRosalesOrtizNo ratings yet
- Ficha Cuantificador Muchos - Pocos (Matemática)Document1 pageFicha Cuantificador Muchos - Pocos (Matemática)TaniaZayuriRosalesOrtizNo ratings yet
- Ficha Cuantificador Muchos - Pocos (Matemática)Document1 pageFicha Cuantificador Muchos - Pocos (Matemática)TaniaZayuriRosalesOrtizNo ratings yet
- Ficha Sonido Inicial de Las Palabras (A-E-I) - (Comunicación)Document1 pageFicha Sonido Inicial de Las Palabras (A-E-I) - (Comunicación)TaniaZayuriRosalesOrtizNo ratings yet
- Secuencia por color y formaDocument6 pagesSecuencia por color y formaTaniaZayuriRosalesOrtizNo ratings yet
- Portada Cuaderno Áreas IntegradasDocument3 pagesPortada Cuaderno Áreas IntegradasTaniaZayuriRosalesOrtizNo ratings yet
- Comunicado Inicio de Talleres - Jardincito de Miss Heidi (4 Años)Document2 pagesComunicado Inicio de Talleres - Jardincito de Miss Heidi (4 Años)TaniaZayuriRosalesOrtizNo ratings yet
- Tarea Repaso Vocales - Descubro Sonido Inicial (Comunicación-Cuaderno Celeste)Document1 pageTarea Repaso Vocales - Descubro Sonido Inicial (Comunicación-Cuaderno Celeste)TaniaZayuriRosalesOrtizNo ratings yet
- Practica 1, Rosales Ortiz - Zamora OrmeñoDocument26 pagesPractica 1, Rosales Ortiz - Zamora OrmeñoTaniaZayuriRosalesOrtizNo ratings yet
- Tarea Número 10 - Formamos Conjuntos y Escritura (Matemática-Cuaderno Amarillo)Document1 pageTarea Número 10 - Formamos Conjuntos y Escritura (Matemática-Cuaderno Amarillo)TaniaZayuriRosalesOrtizNo ratings yet
- Infecciones Resp - AltasDocument55 pagesInfecciones Resp - AltasTaniaZayuriRosalesOrtizNo ratings yet
- Tarea Formato para Dictado de Vocales (Comunicación-Cuaderno Celeste)Document1 pageTarea Formato para Dictado de Vocales (Comunicación-Cuaderno Celeste)TaniaZayuriRosalesOrtizNo ratings yet
- Neumonia Viral SD Coq.Document47 pagesNeumonia Viral SD Coq.TaniaZayuriRosalesOrtizNo ratings yet
- Neumonia Viral SD Coq.Document47 pagesNeumonia Viral SD Coq.TaniaZayuriRosalesOrtizNo ratings yet
- Ficha Grafomotricidad - Trazo SerpenteadoDocument1 pageFicha Grafomotricidad - Trazo SerpenteadoTaniaZayuriRosalesOrtizNo ratings yet
- Neumonia Bacteriana y EmpiemaDocument63 pagesNeumonia Bacteriana y EmpiemaTaniaZayuriRosalesOrtizNo ratings yet
- FiebreDocument33 pagesFiebreTaniaZayuriRosalesOrtizNo ratings yet
- Síndrome bronquial obstrucciónDocument25 pagesSíndrome bronquial obstrucciónTaniaZayuriRosalesOrtizNo ratings yet
- Cuaderno+prácticas+microbiología+enológica 14Document17 pagesCuaderno+prácticas+microbiología+enológica 14Carmen CarmensitaNo ratings yet
- Estallido de RocasDocument7 pagesEstallido de RocasEdwin Elías Mejía ReyesNo ratings yet
- Lab FrutoDocument3 pagesLab FrutoAmy MorejónNo ratings yet
- Bocco (2013) Geografía Ambiental PDFDocument27 pagesBocco (2013) Geografía Ambiental PDFJavier El-Canche AscanioNo ratings yet
- Medio Ambiente y Mi BarrioDocument15 pagesMedio Ambiente y Mi BarrioFernandaRiquelme100% (1)
- Efectos LocalesDocument8 pagesEfectos LocalesjavierurbanoNo ratings yet
- Presentación Proyecto Sociointegrador (Subproductos Del Cerdo-El Estero)Document21 pagesPresentación Proyecto Sociointegrador (Subproductos Del Cerdo-El Estero)Fredrick AlvaradoNo ratings yet
- Barranquilla P.O.T. 2006Document273 pagesBarranquilla P.O.T. 2006Krlos NietoNo ratings yet
- Cuadernillo 2bimestre Naturales Sexto A 12-13Document13 pagesCuadernillo 2bimestre Naturales Sexto A 12-13Gonzalo CastilloNo ratings yet
- Laboratorio Fisica Numero 1Document92 pagesLaboratorio Fisica Numero 1Germán López0% (3)
- Informe Puente de BrooklynDocument24 pagesInforme Puente de BrooklynFabián Ruiz100% (3)
- Diseño de Minas - II UnidadDocument103 pagesDiseño de Minas - II Unidadsimon63No ratings yet
- Manual Sobre El Tae Kwon DoDocument236 pagesManual Sobre El Tae Kwon DoJhordhyEstebanHernandezPerez100% (1)
- CACAODocument49 pagesCACAODavid MV86% (7)
- Crecimiento tornillo tratamientosDocument8 pagesCrecimiento tornillo tratamientoskelly gonzales rojasNo ratings yet
- La Elipse y La Parábola en La FísicaDocument8 pagesLa Elipse y La Parábola en La Físicakiss_lohan14164No ratings yet
- TENSOACTIVOSDocument7 pagesTENSOACTIVOSAnonymous 3xmwtAaiuNo ratings yet
- 73 Planta AldeaDocument2 pages73 Planta AldeaPedroLHernandez0% (1)
- INFORME TECNICO MillerDocument39 pagesINFORME TECNICO MillerJair Esteban EscobedoNo ratings yet
- 10 Cuentos CortosDocument10 pages10 Cuentos CortosErubey CabreraNo ratings yet
- Índices Epidemiológicos en OdontologíaDocument16 pagesÍndices Epidemiológicos en OdontologíaAlejandraNo ratings yet
- Aniones-SALES TERNARIASDocument1 pageAniones-SALES TERNARIASChely MuñozNo ratings yet
- 50 Preguntas Sobre TermitasDocument43 pages50 Preguntas Sobre TermitasRogelioNo ratings yet
- Rios de AmericaDocument4 pagesRios de AmericaAlexis Gonzalez GunnerNo ratings yet
- Criterios estructuración edificios albañileríaDocument48 pagesCriterios estructuración edificios albañileríaAnderson Valverde GarciaNo ratings yet
- Masw SuelosDocument10 pagesMasw SuelosGarry Muñoz NiflaNo ratings yet
- Mapa Mundi RealDocument10 pagesMapa Mundi RealNathalie MartinezNo ratings yet