You might also like
- Physiology of Elasmobranch Fishes: Internal ProcessesFrom EverandPhysiology of Elasmobranch Fishes: Internal ProcessesNo ratings yet
- P2C2 Group 3 Lab Report - Protozoa and PoriferaDocument7 pagesP2C2 Group 3 Lab Report - Protozoa and PoriferaAngela D.No ratings yet
- Lab Activty On OsmoregulationDocument6 pagesLab Activty On OsmoregulationKenneth Myro GarciaNo ratings yet
- Examples of CnidariaDocument5 pagesExamples of CnidariaPeh Jia huiNo ratings yet
- Laporan Lisa Selsai Edit LampranDocument22 pagesLaporan Lisa Selsai Edit LampranAnalianahsNo ratings yet
- GroupNo2 Bauyon LabActivityNo15Document7 pagesGroupNo2 Bauyon LabActivityNo15tel krisNo ratings yet
- Summary, Analysis & Review of Peter Godfrey-Smith’s Other MindsFrom EverandSummary, Analysis & Review of Peter Godfrey-Smith’s Other MindsNo ratings yet
- The Physiology of Digestion in Fish: Environmental Biology of Fishes June 1986Document20 pagesThe Physiology of Digestion in Fish: Environmental Biology of Fishes June 1986Dewi NurhayatiNo ratings yet
- 1 Digestion in Teleost Fishes - 2Document23 pages1 Digestion in Teleost Fishes - 2Evi Nurul IhsanNo ratings yet
- Laboratory Experiment 4Document8 pagesLaboratory Experiment 4auramariefijer1121No ratings yet
- Biology Anotomy VertebrateDocument17 pagesBiology Anotomy VertebrateAhmad ZuhudyNo ratings yet
- Anatomy 1 Nilem and Cat FishDocument15 pagesAnatomy 1 Nilem and Cat FishAnisatul Khabibah ZaenNo ratings yet
- Observations of Gut Contents of Leptocephali in THDocument13 pagesObservations of Gut Contents of Leptocephali in THsonia bhanNo ratings yet
- (The Natural History of The Crustacea) Les Watling, Martin Thiel (Eds.) - Functional Morphology and Diversity. 1-Oxford University Press (2013)Document515 pages(The Natural History of The Crustacea) Les Watling, Martin Thiel (Eds.) - Functional Morphology and Diversity. 1-Oxford University Press (2013)Yamaly Barragán MarínNo ratings yet
- Lab 9 - Vertebrate Organ Systems: Biology 18 Spring 2008Document43 pagesLab 9 - Vertebrate Organ Systems: Biology 18 Spring 2008JulsNo ratings yet
- Project ThesisDocument29 pagesProject ThesisImmanuvelNo ratings yet
- Arthropoda Characteristics: Circulatory SystemDocument15 pagesArthropoda Characteristics: Circulatory SystemBello AjetayoNo ratings yet
- BIO 85 Vertebral Column AssignmentDocument5 pagesBIO 85 Vertebral Column AssignmentvinlavisoNo ratings yet
- Physiology of Elasmobranch Fishes: Structure and Interaction with EnvironmentFrom EverandPhysiology of Elasmobranch Fishes: Structure and Interaction with EnvironmentNo ratings yet
- Aqu247 Lab Report 2 Syafiz Iqmal 2019412616Document9 pagesAqu247 Lab Report 2 Syafiz Iqmal 2019412616SYAFIZ IQMAL RAMLANNo ratings yet
- ZF and CAVEFISHDocument6 pagesZF and CAVEFISHsultanmsajidmNo ratings yet
- Frog DissectionDocument2 pagesFrog DissectionKylle CalipesNo ratings yet
- ProtistDocument5 pagesProtistGerald Agacid BangeroNo ratings yet
- Laporan KatakDocument17 pagesLaporan Katakbertha tandiNo ratings yet
- Neurobiology and BehaviorFrom EverandNeurobiology and BehaviorA. O. Dennis WillowsNo ratings yet
- Biotechnological Potential of Marine Sponges: Narsinh L. Thakur and Werner E. G. MüllerDocument7 pagesBiotechnological Potential of Marine Sponges: Narsinh L. Thakur and Werner E. G. MüllerIbnu AriyantoNo ratings yet
- CordovillaMark King Joseph P - MODULE8 - ZOOLOGYDocument2 pagesCordovillaMark King Joseph P - MODULE8 - ZOOLOGYMark king Joseph CordovillaNo ratings yet
- Zooplankton ReportDocument6 pagesZooplankton Reportapi-252723480No ratings yet
- Electron Microscopy of The Intestinal Microflora of FishDocument21 pagesElectron Microscopy of The Intestinal Microflora of FishRima Oktaviani PutriNo ratings yet
- Virtual Laboratory: Organisms in Pond Water: Group Number - Members Galero - Guotana - Leonor - NebrejaDocument4 pagesVirtual Laboratory: Organisms in Pond Water: Group Number - Members Galero - Guotana - Leonor - NebrejaHedda Mae LeonorNo ratings yet
- Lab 6 - Survey of MicroorganismDocument6 pagesLab 6 - Survey of MicroorganismYusof SundangNo ratings yet
- Zelfstudie Van Blastula Naar 3 KiemlagenDocument18 pagesZelfstudie Van Blastula Naar 3 KiemlagenCheyenne ProvoostNo ratings yet
- Porifera: Sponge (Tool) Sponge (Disambiguation)Document5 pagesPorifera: Sponge (Tool) Sponge (Disambiguation)zoology qauNo ratings yet
- Biology 11 Unit 9 Assignment 1 How Do Sponges Cnidarians Flatworms and Roundworms Obtain Food Virtual LabDocument4 pagesBiology 11 Unit 9 Assignment 1 How Do Sponges Cnidarians Flatworms and Roundworms Obtain Food Virtual Labapi-323453767No ratings yet
- Lab 5 Mollusks Write UpDocument4 pagesLab 5 Mollusks Write Upm.iabobeuhNo ratings yet
- Laboratory Dissection - Phylum Mollusca FinalDocument8 pagesLaboratory Dissection - Phylum Mollusca Finalapi-224938251No ratings yet
- Free AIPMT BIOLOGY Study MaterialDocument346 pagesFree AIPMT BIOLOGY Study MaterialApex Iit-jee87% (54)
- A Treatise on Physiology and Hygiene: For Educational Institutions and General ReadersFrom EverandA Treatise on Physiology and Hygiene: For Educational Institutions and General ReadersNo ratings yet
- Chick EmbryologyDocument55 pagesChick EmbryologySushma MishraNo ratings yet
- Koleksi SpesimenDocument8 pagesKoleksi SpesimenDaisy KavinskyNo ratings yet
- Phagocytic - Role in DigestionDocument4 pagesPhagocytic - Role in Digestionchardz10No ratings yet
- Laporan Anatomi Ikan Nilem Dan Ikan LeleDocument9 pagesLaporan Anatomi Ikan Nilem Dan Ikan LeleDaisy KavinskyNo ratings yet
- Crayfish Dissection LabDocument2 pagesCrayfish Dissection LabKate FairleyNo ratings yet
- Laprak SisheewDocument8 pagesLaprak SisheewMellya RizkiNo ratings yet
- The Biology and Management of Lobsters: Ecology and ManagementFrom EverandThe Biology and Management of Lobsters: Ecology and ManagementJ. Stanley CobbNo ratings yet
- Laprak IkanDocument13 pagesLaprak IkanRezky AmaliahNo ratings yet
- Biology - Pond Water LabDocument4 pagesBiology - Pond Water LabAngela CuiNo ratings yet
- BZ Lab 5Document8 pagesBZ Lab 5Alexa Jean D. HonrejasNo ratings yet
- AIPMT BIOLOGY Study Material PDFDocument346 pagesAIPMT BIOLOGY Study Material PDFDanish BoddaNo ratings yet
- 1 Cephalopod Biology and Ongoing ResearchDocument9 pages1 Cephalopod Biology and Ongoing ResearchVicente VasquezNo ratings yet
- The Evidence: Ch. 2 Tiktaalik/ The Rotation of Our Knees and Elbows in UteroDocument13 pagesThe Evidence: Ch. 2 Tiktaalik/ The Rotation of Our Knees and Elbows in UteroCory LukerNo ratings yet
- Complete Report Unit 4Document24 pagesComplete Report Unit 4Nisa AndiniNo ratings yet
- Histology of Umbilical Cord in MammalsDocument16 pagesHistology of Umbilical Cord in MammalsFia NisaNo ratings yet
- Exercise 8 PlacentaDocument6 pagesExercise 8 PlacentaPrabhatNo ratings yet
- Fan Tool Kit - Ad Hoc Group - V4ddDocument121 pagesFan Tool Kit - Ad Hoc Group - V4ddhmaza shakeelNo ratings yet
- API 16C ErrataDocument1 pageAPI 16C ErrataDinesh KumarNo ratings yet
- ISKCON Desire Tree - Brahma Vimohana LeelaDocument34 pagesISKCON Desire Tree - Brahma Vimohana LeelaISKCON desire treeNo ratings yet
- PiscicultureDocument14 pagesPisciculturejisha26100% (1)
- Static Fluid: It Is Mass Per Unit Volume at Certain TemperatureDocument11 pagesStatic Fluid: It Is Mass Per Unit Volume at Certain TemperatureMohamed El-GoharyNo ratings yet
- Entomotherapy, or The Medicinal Use of InsectsDocument23 pagesEntomotherapy, or The Medicinal Use of InsectsJessica Revelo OrdoñezNo ratings yet
- Flight School BundleDocument43 pagesFlight School BundleyoyoNo ratings yet
- ISO 9001 Required DocumentationDocument3 pagesISO 9001 Required Documentationdnmule100% (1)
- The Emerging Field of Emotion Regulation - An Integrative Review - James J GrossDocument29 pagesThe Emerging Field of Emotion Regulation - An Integrative Review - James J GrossLely AprianiNo ratings yet
- MUMMY'S KITCHEN NEW FinalDocument44 pagesMUMMY'S KITCHEN NEW Finalanon_602671575100% (4)
- Method Setting Out of Drainage 2Document9 pagesMethod Setting Out of Drainage 2amin100% (1)
- ACRE2a Non Elementary Reaction Kinetics RevDocument53 pagesACRE2a Non Elementary Reaction Kinetics RevRathish RagooNo ratings yet
- AcuityPDR BrochureDocument3 pagesAcuityPDR BrochureJulian HutabaratNo ratings yet
- 1erTareaMicroscopíasLópez Marmolejo Clere MishellDocument4 pages1erTareaMicroscopíasLópez Marmolejo Clere Mishellclere02marmolejoNo ratings yet
- IFAD Vietnam RIMS Training Workshop 2011 (1 of 7)Document18 pagesIFAD Vietnam RIMS Training Workshop 2011 (1 of 7)IFAD VietnamNo ratings yet
- DemolitionDocument25 pagesDemolitionusler4u100% (1)
- Scala: Service InstructionsDocument16 pagesScala: Service Instructionsmario_turbinadoNo ratings yet
- Neicchi 270 ManualDocument33 pagesNeicchi 270 Manualmits2004No ratings yet
- 10 Major Sulfuric Acid Industrial Applications - WorldOfChemicalsDocument9 pages10 Major Sulfuric Acid Industrial Applications - WorldOfChemicalsFarhad MalikNo ratings yet
- Soil ExplorationDocument42 pagesSoil ExplorationDilipKumarAkkaladevi100% (1)
- Grade 10 Chemistry Week 1Document3 pagesGrade 10 Chemistry Week 1Nikoli MajorNo ratings yet
- Behavior Modification: Presented By: Wajeeha Iqbal BS in Psychology, MS (SLP/T) Riphah College of Rehabilitation SciencesDocument28 pagesBehavior Modification: Presented By: Wajeeha Iqbal BS in Psychology, MS (SLP/T) Riphah College of Rehabilitation Sciencesmazahir hussainNo ratings yet
- 01 Head and Neck JMDocument16 pages01 Head and Neck JMJowi SalNo ratings yet
- CS1 Epoxy Curing AgentsDocument21 pagesCS1 Epoxy Curing AgentsAlptekin100% (1)
- Lecture 38Document10 pagesLecture 38Deepak GuptaNo ratings yet
- ND 0108 SpongeDiver SymiDocument2 pagesND 0108 SpongeDiver SymiPoki MokiNo ratings yet
- EN Ultra V Mini STP410S C54 UmhDocument2 pagesEN Ultra V Mini STP410S C54 Umhjabt4568No ratings yet
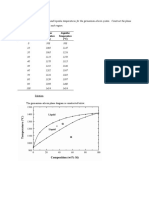
- Composition Solidus Temperature Liquidus Temperature: (WT% Si) (°C) (°C)Document7 pagesComposition Solidus Temperature Liquidus Temperature: (WT% Si) (°C) (°C)Muhammad Ibkar YusranNo ratings yet
- Complications of Diabetes MellitusDocument46 pagesComplications of Diabetes MellitusAbhijith MenonNo ratings yet
- Tutorials 2016Document54 pagesTutorials 2016Mankush Jain100% (1)