You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5783)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- How Sleep Deprivation Affects Students' Academic PerformanceDocument50 pagesHow Sleep Deprivation Affects Students' Academic PerformanceAstrophile0% (2)
- Lefort 1 OsteotomyDocument16 pagesLefort 1 Osteotomyabhiram.ramesh1290No ratings yet
- Who Needs Bypass SurgeryDocument70 pagesWho Needs Bypass SurgerymikikiNo ratings yet
- Parasitology Lect. (4) : Preparation of Solutions For Wet MountDocument5 pagesParasitology Lect. (4) : Preparation of Solutions For Wet MountAbdul hadiNo ratings yet
- Globin Synthesis PDFDocument6 pagesGlobin Synthesis PDFmanoj_rkl_07100% (1)
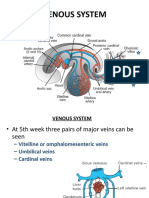
- Development - of - CVS-5Document20 pagesDevelopment - of - CVS-5Ahsan IslamNo ratings yet
- Prokaryotic Cells: Eukaryotes Vs Prokaryotes: What's The Difference?Document8 pagesProkaryotic Cells: Eukaryotes Vs Prokaryotes: What's The Difference?Anggun Teh PamegetNo ratings yet
- Autopsy Report: DECEDENT: John AndersonDocument5 pagesAutopsy Report: DECEDENT: John AndersonjilliankadNo ratings yet
- The Circulatory SystemDocument8 pagesThe Circulatory Systemrosario marcelaNo ratings yet
- Ayurveda Physical BodyDocument11 pagesAyurveda Physical BodyanantNo ratings yet
- Teachers' Health Examination Card (CS Form 86)Document4 pagesTeachers' Health Examination Card (CS Form 86)Maria Angellie S. Bellido - EramisNo ratings yet
- Laboratory Exercise No.8Document3 pagesLaboratory Exercise No.8yumi castroNo ratings yet
- ANS & CVS Response To Exercise v2Document18 pagesANS & CVS Response To Exercise v2chow wing yin amandaNo ratings yet
- Defense Mechanisms of the GingivaDocument249 pagesDefense Mechanisms of the GingivaAssssssNo ratings yet
- Receptors IntroductionDocument53 pagesReceptors IntroductionSunilNo ratings yet
- Hyperlipedemia PPDocument51 pagesHyperlipedemia PPrabarNo ratings yet
- Guillain Barre SyndromeDocument2 pagesGuillain Barre SyndromeTheresia Avila KurniaNo ratings yet
- Newborn Hypothermia and Hyperthermia (Final Copy)Document30 pagesNewborn Hypothermia and Hyperthermia (Final Copy)Shizuka Marycris Amane67% (3)
- The Human Body in Health and Illness e Book 6th Edition Ebook PDFDocument61 pagesThe Human Body in Health and Illness e Book 6th Edition Ebook PDFcraig.suarez45797% (33)
- Anhidrosis in EquineDocument3 pagesAnhidrosis in EquineZain AsifNo ratings yet
- HBCJ Npev Vedw T60B PDFDocument20 pagesHBCJ Npev Vedw T60B PDFAnonymous fPQaCe8No ratings yet
- Histological Characteristics of the Lingual FrenulumDocument1 pageHistological Characteristics of the Lingual FrenulumajudNo ratings yet
- MEBO Moisture and Wound HealingDocument6 pagesMEBO Moisture and Wound HealingAnonymous 8hVpaQdCtr100% (2)
- Motility Disorders of EsophagusDocument44 pagesMotility Disorders of EsophagusmackieccNo ratings yet
- Peripheral Testosterone MetabolismDocument8 pagesPeripheral Testosterone MetabolismAgung SentosaNo ratings yet
- Pain Aapa Rev8Document101 pagesPain Aapa Rev8Novii NoviiNo ratings yet
- Autonomic Nervous System For MBBSDocument20 pagesAutonomic Nervous System For MBBSjacobsNo ratings yet
- 76) Book Chapter 2010Document11 pages76) Book Chapter 2010Giovanni MauroNo ratings yet
- Bioactive Lipid Mediators - (2015)Document424 pagesBioactive Lipid Mediators - (2015)MohammedAjebliNo ratings yet
- Figure 1 Human Nervous System Source:courses - Lumenlearning.c Om/microbiology/chapter/anato My-Of-The-Nervous-SystemDocument14 pagesFigure 1 Human Nervous System Source:courses - Lumenlearning.c Om/microbiology/chapter/anato My-Of-The-Nervous-SystemShekaina Faith Cuizon LozadaNo ratings yet