You might also like
- Metabolic Concepts (Lecture 1-3)Document8 pagesMetabolic Concepts (Lecture 1-3)atifzea100% (1)
- Lecture - 3 Cell Metabolism: Dr. S. Annie Jeyachristy Lecturer, Unit of Biochemistry Faculty of MedicineDocument15 pagesLecture - 3 Cell Metabolism: Dr. S. Annie Jeyachristy Lecturer, Unit of Biochemistry Faculty of MedicinecheckmateNo ratings yet
- Biochemistry PDFDocument114 pagesBiochemistry PDFShanmuga Moorthy100% (2)
- Lecture Two: Carbohydrate Lecture Two: CarbohydrateDocument24 pagesLecture Two: Carbohydrate Lecture Two: Carbohydratesaacid bashir100% (1)
- MCQ Biochemistry CarbohydrateDocument21 pagesMCQ Biochemistry Carbohydratestuff100% (1)
- BiochemistryDocument82 pagesBiochemistryAmritzz Paul83% (6)
- MCQs PDFDocument307 pagesMCQs PDFChirag Dagar50% (2)
- Digestion and Absorption of LipidsDocument5 pagesDigestion and Absorption of LipidsAngelo Jude Cobacha100% (1)
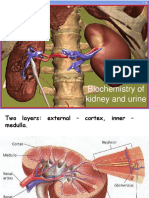
- Biochemistry of Kidney and UrineDocument19 pagesBiochemistry of Kidney and UrineKarina PathyaNo ratings yet
- Heavy Metal PoisoningDocument15 pagesHeavy Metal PoisoningfianceeleeNo ratings yet
- CELLDocument45 pagesCELLKimberly Clarisse VegaNo ratings yet
- Chapter 1 AnaPhy Notes - ReviewerDocument6 pagesChapter 1 AnaPhy Notes - ReviewerDanielle LeighNo ratings yet
- Genetic Engineering (Gowshia, Fatimah, Inthuja)Document15 pagesGenetic Engineering (Gowshia, Fatimah, Inthuja)Lacramioara Amalia Amariei100% (1)
- HomeostasisDocument16 pagesHomeostasisMerly MorgianaNo ratings yet
- Biochemistry of Kidneys and UrineDocument18 pagesBiochemistry of Kidneys and UrineAndrias PutriNo ratings yet
- 6.1 Types of NutritionDocument11 pages6.1 Types of NutritionNoor Hidayah SambliNo ratings yet
- 1-Biochemistry Chapter No 1Document63 pages1-Biochemistry Chapter No 1Mahrukh Saeed100% (1)
- 4 19 Lymphmatic SystemDocument10 pages4 19 Lymphmatic SystemJohn Lawrence Ybanez100% (1)
- 1 - XenobioticsDocument24 pages1 - XenobioticsgeenaksamuelNo ratings yet
- Steatosis and SteatohapatitisDocument47 pagesSteatosis and SteatohapatitisFifi RohmatinNo ratings yet
- Human Anatomy and PhysiologyDocument46 pagesHuman Anatomy and PhysiologyBegumHazinNo ratings yet
- Pre-Finals Topic 1 AnaphyDocument5 pagesPre-Finals Topic 1 AnaphyGlenice Joy SenocNo ratings yet
- The Origin and Chemistry of Life: Prepared By: Asst. Prof. Sheryl Santa Cruz-BiscochoDocument25 pagesThe Origin and Chemistry of Life: Prepared By: Asst. Prof. Sheryl Santa Cruz-Biscochoirish x100% (1)
- Urea CycleDocument11 pagesUrea CycleRohit VinayNo ratings yet
- The Digestive SystemDocument30 pagesThe Digestive SystemUniversal DiscoveringNo ratings yet
- Acfrogas81a C3gdggyv4xgrao Mdfic3te - x187f1 87lztljhozqte1uwyasvd30we4h6qfxqnls Kkx4fhvgdzsyhiidv6ruak2jsebl1ta-o6xhpricpgy55ribskr1dafecu67-RbikzqhDocument126 pagesAcfrogas81a C3gdggyv4xgrao Mdfic3te - x187f1 87lztljhozqte1uwyasvd30we4h6qfxqnls Kkx4fhvgdzsyhiidv6ruak2jsebl1ta-o6xhpricpgy55ribskr1dafecu67-RbikzqhSiseneg Tasma100% (1)
- Diet NutritionDocument37 pagesDiet NutritionVikesh SharmaNo ratings yet
- BIOENERGETICS - How The Body Converts Food To Energy - Group 7 (MC 102 - Lecture) EDITEDDocument73 pagesBIOENERGETICS - How The Body Converts Food To Energy - Group 7 (MC 102 - Lecture) EDITEDJowe VarnalNo ratings yet
- Unit 4 Factors Affecting Drug AbsorptionDocument53 pagesUnit 4 Factors Affecting Drug Absorptiondiadiaida100% (1)
- Metabolism QuestionsDocument5 pagesMetabolism QuestionsErjus Hoxhaj100% (1)
- Lipid MetabolismDocument17 pagesLipid MetabolismMiran El-MaghrabiNo ratings yet
- HomeostasisDocument35 pagesHomeostasisPranshu AdityaNo ratings yet
- Dna and Rna Powerpoint 2Document46 pagesDna and Rna Powerpoint 2api-267309851No ratings yet
- Ecological RelationshipsDocument31 pagesEcological RelationshipsLiliana ŞeremetNo ratings yet
- Biology NotesDocument6 pagesBiology NotesElizaNo ratings yet
- Intellectual RevolutionDocument13 pagesIntellectual Revolutionblueviolet21100% (1)
- CH 03 Lecture Presentation-1 PDFDocument136 pagesCH 03 Lecture Presentation-1 PDFanon_33138328100% (1)
- Chapter 2 - Basics of EnzymesDocument40 pagesChapter 2 - Basics of EnzymesSakinah MuhamadNo ratings yet
- Frog S Muscular System PDFDocument9 pagesFrog S Muscular System PDFVia CabardaNo ratings yet
- Biochemistry MCQ, Biochem by PQP, WWW - Pharmaquizportal.tk, Pharma Quiz Point & Pharma Quiz PortalDocument10 pagesBiochemistry MCQ, Biochem by PQP, WWW - Pharmaquizportal.tk, Pharma Quiz Point & Pharma Quiz PortalDrug Viral100% (1)
- Notes For HomeostasisDocument21 pagesNotes For Homeostasisfreednemon83% (6)
- Complexation and Protein BindingDocument52 pagesComplexation and Protein BindingPankaj Patel100% (1)
- Enzymes and Their Importance in Plants and AnimalsDocument4 pagesEnzymes and Their Importance in Plants and Animalsanili50% (2)
- Diff Osmosis Lab Sp11Document8 pagesDiff Osmosis Lab Sp11Kelly TrainorNo ratings yet
- Glucose Homeostasis: by Dr. Sumbul FatmaDocument28 pagesGlucose Homeostasis: by Dr. Sumbul Fatmaraanja2No ratings yet
- Digestive System NotesDocument10 pagesDigestive System NotesArchanna VyassNo ratings yet
- Bioavailability: Factor AffectingDocument11 pagesBioavailability: Factor AffectingMalvinder SharmaNo ratings yet
- Chromatographic Separation of Amino AcidsDocument1 pageChromatographic Separation of Amino AcidsManP13No ratings yet
- Reviewer ANPH111 CandyDocument21 pagesReviewer ANPH111 CandyRein TopacioNo ratings yet
- Ribosomes and Protein SynthesisDocument12 pagesRibosomes and Protein SynthesisSulfeekker AhamedNo ratings yet
- Chapter I Structural OrganizationDocument15 pagesChapter I Structural OrganizationTitoMacoyTVNo ratings yet
- BPH First Year Curriculum of Purbanchal UniversityDocument48 pagesBPH First Year Curriculum of Purbanchal Universityseo167No ratings yet
- 2013 Body Systems NotesDocument5 pages2013 Body Systems Notesapi-327757745No ratings yet
- Review On THE Bioavailability of Phytochemicals and Their Role in Nutrition and HealthDocument32 pagesReview On THE Bioavailability of Phytochemicals and Their Role in Nutrition and HealthAbraha AbadiNo ratings yet
- Biochemistry: DR - Radhwan M. Asal Bsc. Pharmacy MSC, PHD Clinical BiochemistryDocument13 pagesBiochemistry: DR - Radhwan M. Asal Bsc. Pharmacy MSC, PHD Clinical BiochemistryAnas SeghayerNo ratings yet
- Denaturation of ProteinsDocument32 pagesDenaturation of ProteinsAhmad KhanNo ratings yet
- SubcellularFractionation Fa15Document25 pagesSubcellularFractionation Fa15Duane HallNo ratings yet
- Body System ChecklistDocument6 pagesBody System Checklistapi-422967453No ratings yet
- A4 QB-MC Ch02 The Cell As The Basic Unit of LifeDocument29 pagesA4 QB-MC Ch02 The Cell As The Basic Unit of LifeReg ChooNo ratings yet
- High School Science Virtual Learning: Biology Homeostasi SDocument20 pagesHigh School Science Virtual Learning: Biology Homeostasi SEdchel EspeñaNo ratings yet
- Inorganic and Organometallic Reaction MechanismsDocument15 pagesInorganic and Organometallic Reaction MechanismsKartikeya Singh0% (1)
- Tec No StudiesDocument34 pagesTec No StudiesarthurhuntNo ratings yet
- Microbiology With Diseases by Taxonomy 5th Edition Bauman Solutions Manual 1Document36 pagesMicrobiology With Diseases by Taxonomy 5th Edition Bauman Solutions Manual 1tinabrooksidzrkgytcx100% (23)
- BioN06 Generation of Biochemical Energy Summer 2015Document69 pagesBioN06 Generation of Biochemical Energy Summer 2015glenn floresNo ratings yet
- Genomic Analysis of MAP Kinase Cascades In: Arabidopsis Defense ResponsesDocument13 pagesGenomic Analysis of MAP Kinase Cascades In: Arabidopsis Defense ResponsesMelvin PrasadNo ratings yet
- GlucagonDocument4 pagesGlucagonElenanana100% (1)
- BCH GlycolysisDocument14 pagesBCH GlycolysisscholasticaNo ratings yet
- Glycogen Metabolism - Index CardDocument3 pagesGlycogen Metabolism - Index CardKate Alyssa CatonNo ratings yet
- Fundamentals of Glycogen.Document17 pagesFundamentals of Glycogen.Kevin VillaNo ratings yet
- Janes Metro Map Pathways BLANKDocument1 pageJanes Metro Map Pathways BLANKWinston TengNo ratings yet
- Notes+4 +ATP,+Water+and+Inorganic+IonsDocument5 pagesNotes+4 +ATP,+Water+and+Inorganic+IonsSyeda Wardah NoorNo ratings yet
- Carbohydrate Metabolism BioChem Lec Finals2x1 FormatDocument3 pagesCarbohydrate Metabolism BioChem Lec Finals2x1 FormatSheene ImblarinagNo ratings yet
- Fatty Acid Synthesis: Molecular Biochemistry IIDocument31 pagesFatty Acid Synthesis: Molecular Biochemistry IIDozdiNo ratings yet
- Diabetes Case Study - Jason BaoDocument8 pagesDiabetes Case Study - Jason Baoapi-522847737No ratings yet
- Module 5 CarbohydratesDocument21 pagesModule 5 CarbohydratesSpongebob SquarepantsNo ratings yet
- Glycolysis PathwayDocument29 pagesGlycolysis PathwayAinsleyNo ratings yet
- Hematology 1 MidtermsDocument31 pagesHematology 1 Midtermsella SyNo ratings yet
- CC 11 3611Document16 pagesCC 11 3611Sergeat18BNo ratings yet
- MOL 16 - Intro To MetabolismDocument8 pagesMOL 16 - Intro To Metabolismqueenmasa191No ratings yet
- Insulin-Induced Hexokinase II Expression Is Reduced in Obesity and NIDDMDocument8 pagesInsulin-Induced Hexokinase II Expression Is Reduced in Obesity and NIDDMUmarani MohanasundaramNo ratings yet
- Chapter 15 SummaryDocument27 pagesChapter 15 SummaryCharlotteNo ratings yet
- I. A New Diet: Table 1.the Biological Effects of InsulinDocument9 pagesI. A New Diet: Table 1.the Biological Effects of InsulinmelisaNo ratings yet
- Biology 1113 Notes Unit 4Document17 pagesBiology 1113 Notes Unit 4Shane YacyshynNo ratings yet
- Xu Princeton 0181D 10615Document175 pagesXu Princeton 0181D 10615Sailendra MeherNo ratings yet
- Glycolysis, Glycogenesis, Glycogenolysis and GluconeogensisDocument103 pagesGlycolysis, Glycogenesis, Glycogenolysis and GluconeogensisHabibur RahmanNo ratings yet
- Human Physiology 13Th Edition Stuart Ira Fox Test Bank Full Chapter PDFDocument36 pagesHuman Physiology 13Th Edition Stuart Ira Fox Test Bank Full Chapter PDFevelyn.trudnowski238100% (14)
- Cell Structure and Funtions 2 PDFDocument49 pagesCell Structure and Funtions 2 PDFEllie MarrisNo ratings yet
- Nikhil BTP Mid Term ReportDocument17 pagesNikhil BTP Mid Term ReportNikhil SaiyamNo ratings yet