You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- 8 Causes of Periumbilical Pain and When To Seek Emergency HelpDocument8 pages8 Causes of Periumbilical Pain and When To Seek Emergency HelpJohn Rodney PicaNo ratings yet
- Acute CholecystitisDocument6 pagesAcute Cholecystitisrini9384No ratings yet
- Approach To Patients With Abdominal Pain-ValenDocument49 pagesApproach To Patients With Abdominal Pain-ValenCitra Sucipta0% (1)
- Pointer Trial Long Term FinalDocument43 pagesPointer Trial Long Term FinalHarsha VardhanNo ratings yet
- Acute PancreatitisDocument9 pagesAcute PancreatitisJayari Cendana PutraNo ratings yet
- Broken Rib SymptomsDocument12 pagesBroken Rib Symptomsneleh grayNo ratings yet
- M Digestion" Absorption" EliminationDocument104 pagesM Digestion" Absorption" EliminationChristine Carol FilipinasNo ratings yet
- A Lexicon For Endoscopic Adverse Events: Report of An ASGE WorkshopDocument9 pagesA Lexicon For Endoscopic Adverse Events: Report of An ASGE WorkshopResci Angelli Rizada-NolascoNo ratings yet
- TVP-2020-0304 Acute PancreatitisDocument7 pagesTVP-2020-0304 Acute Pancreatitischeldon jacobNo ratings yet
- Chronic PancreatitisDocument2 pagesChronic PancreatitisIbraheem SairiNo ratings yet
- PancreatitisDocument7 pagesPancreatitisavigenNo ratings yet
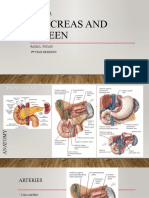
- Pancreas and SpleenDocument106 pagesPancreas and SpleenJorge De Vera100% (1)
- Seminar: Definition of Chronic PancreatitisDocument14 pagesSeminar: Definition of Chronic PancreatitisCarlos Alberto Torres LópezNo ratings yet
- Case 13 Medical Nutrition TherapyDocument8 pagesCase 13 Medical Nutrition TherapykleeNo ratings yet
- Acute Pancreatitis Pathophysiology Clinical AspectDocument6 pagesAcute Pancreatitis Pathophysiology Clinical AspectTrajceNo ratings yet
- Section D - Work BookDocument69 pagesSection D - Work BookMohamed AbbasNo ratings yet
- Surgery Cycle MCQSDocument27 pagesSurgery Cycle MCQSHeeb Warda100% (1)
- MOCKBOARD PART 3 Answer KeyDocument23 pagesMOCKBOARD PART 3 Answer KeyJayrald CruzadaNo ratings yet
- 12 Health Risks of Chronic Heavy DrinkingDocument3 pages12 Health Risks of Chronic Heavy Drinkingcyberbat2008No ratings yet
- Pancreatic PseudocystDocument3 pagesPancreatic PseudocystViorel BudacNo ratings yet
- Acute PancreatitisDocument4 pagesAcute Pancreatitissmith dNo ratings yet
- Chronic Pancreatitis: PathophysiologyDocument7 pagesChronic Pancreatitis: PathophysiologyMadelaine De Arco J.No ratings yet
- Approach To Acute AbdomenDocument50 pagesApproach To Acute AbdomenAndie ArrNo ratings yet
- Pancreatitis ChronicDocument9 pagesPancreatitis ChronicGherca IzabellaNo ratings yet
- Case Study Cirrhosis of LiverDocument26 pagesCase Study Cirrhosis of LiverMonika Seervi100% (1)
- Nausea and Vomiting in Adolescents and AdultsDocument30 pagesNausea and Vomiting in Adolescents and AdultsPramita Ines ParmawatiNo ratings yet
- Plabable Gems Gastroenterology PDF h6sDocument39 pagesPlabable Gems Gastroenterology PDF h6sayşenur sevinçNo ratings yet
- Chronic Pancreatitis (LitReview - 2009)Document28 pagesChronic Pancreatitis (LitReview - 2009)Dimsy Flora KpaiNo ratings yet
- Sklera Sub IkterikDocument7 pagesSklera Sub IkterikFauziah_Hannum_SNo ratings yet
- Week2Handout - NDT - Liver, Gallbladder, and PancreasDocument7 pagesWeek2Handout - NDT - Liver, Gallbladder, and PancreasMegumi S.No ratings yet