You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Equine Reproductive Physiology, Breeding, and Stud ManagementDocument383 pagesEquine Reproductive Physiology, Breeding, and Stud Managementfrancisco_aguiar_22100% (2)
- Applications of MatricesDocument45 pagesApplications of MatricesYashwanth_Hari_5831100% (1)
- Shil and Debnath Part 2Document1 pageShil and Debnath Part 2rafiqcuNo ratings yet
- KhatianDocument3 pagesKhatianrafiqcuNo ratings yet
- NTPC Harming Bangladesh's Sundarbans' - Dhaka TribuneDocument1 pageNTPC Harming Bangladesh's Sundarbans' - Dhaka TribunerafiqcuNo ratings yet
- Excursion To Sempu IslandDocument2 pagesExcursion To Sempu IslandrafiqcuNo ratings yet
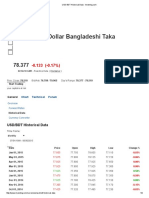
- USD BDT Historical Data - InvestingDocument9 pagesUSD BDT Historical Data - InvestingrafiqcuNo ratings yet
- Prayer Time Chittagong 2016Document3 pagesPrayer Time Chittagong 2016rafiqcuNo ratings yet
- Cramers Rule ProofDocument10 pagesCramers Rule ProofrafiqcuNo ratings yet
- Tirish Bochor Pore o Shadinoa Ta Ke KujchiDocument3 pagesTirish Bochor Pore o Shadinoa Ta Ke KujchirafiqcuNo ratings yet
- The Best Preparation For IELTS Writing PDFDocument251 pagesThe Best Preparation For IELTS Writing PDFrafiqcu91% (11)
- Electronic Green JournalDocument23 pagesElectronic Green JournalrafiqcuNo ratings yet
- Proceedings of The International Conference On Biodiversity CU Raihan and Rosket CoverDocument1 pageProceedings of The International Conference On Biodiversity CU Raihan and Rosket CoverrafiqcuNo ratings yet
- Tokyo Tech Muslim GuideDocument20 pagesTokyo Tech Muslim GuiderafiqcuNo ratings yet
- Easy Calculation SiteDocument2 pagesEasy Calculation SiterafiqcuNo ratings yet
- Bumpy Start For Malaysia's First Sharia-Compliant Airline - The DiplomatDocument2 pagesBumpy Start For Malaysia's First Sharia-Compliant Airline - The DiplomatrafiqcuNo ratings yet
- Marine Resource Economics - HomeDocument3 pagesMarine Resource Economics - HomerafiqcuNo ratings yet
- Will This Weapon Change Infantry Warfare Forever - The DiplomatDocument3 pagesWill This Weapon Change Infantry Warfare Forever - The DiplomatrafiqcuNo ratings yet
- The State of Lie-Flat Business Class On American CarriersThe Points GuyDocument12 pagesThe State of Lie-Flat Business Class On American CarriersThe Points GuyrafiqcuNo ratings yet
- Uzbekistan - White Gold, Dirty Business - The DiplomatDocument3 pagesUzbekistan - White Gold, Dirty Business - The DiplomatrafiqcuNo ratings yet
- Japan's Achilles Heel - Cybersecurity - The DiplomatDocument5 pagesJapan's Achilles Heel - Cybersecurity - The DiplomatrafiqcuNo ratings yet
- Avoid PlagiarismDocument16 pagesAvoid PlagiarismrafiqcuNo ratings yet
- Trees in QuranDocument8 pagesTrees in QuranrafiqcuNo ratings yet
- Reference Paper and My Title For PresentationDocument67 pagesReference Paper and My Title For PresentationrafiqcuNo ratings yet
- Carbon CrotonDocument2 pagesCarbon CrotonrafiqcuNo ratings yet
- Okinawa Palying Role in Mangrove ProtectionDocument1 pageOkinawa Palying Role in Mangrove ProtectionrafiqcuNo ratings yet
- PNAS 2010 Mori 1447 51Document5 pagesPNAS 2010 Mori 1447 51rafiqcuNo ratings yet
- Journal of Political EcologyDocument12 pagesJournal of Political EcologySerdar BıcakcıNo ratings yet
- Lessons Learnt From Buchanan BookDocument1 pageLessons Learnt From Buchanan BookrafiqcuNo ratings yet
- Bibliography Teaching For DepthDocument49 pagesBibliography Teaching For DepthrafiqcuNo ratings yet
- DR ScrapbookDocument8 pagesDR Scrapbookcrenz23_shaniyha15No ratings yet
- Lesson Plan For Final DemoDocument9 pagesLesson Plan For Final DemoJunjun Caoli50% (2)
- PlacentaDocument6 pagesPlacentaKishore Singh BasnetNo ratings yet
- The Miracle of Life, Images of Gestation of Baby in The Womb, in Utero Photos of An Embryo With Development From Conception To Birth.Document17 pagesThe Miracle of Life, Images of Gestation of Baby in The Womb, in Utero Photos of An Embryo With Development From Conception To Birth.Lily of the ValleysNo ratings yet
- Introduction To Life Science, Earth and LifeDocument16 pagesIntroduction To Life Science, Earth and Lifejoel TorresNo ratings yet
- The REPRODUCTION-PROCESSDocument30 pagesThe REPRODUCTION-PROCESSEJ GolimlimNo ratings yet
- Bentham and Hooker Classification System - OecDocument22 pagesBentham and Hooker Classification System - OecDAVALDARIYA WALIKARNo ratings yet
- Science QuizDocument13 pagesScience Quizgrethel anne abellanosaNo ratings yet
- Algae 2022 37 5 11Document17 pagesAlgae 2022 37 5 11Kathya Gómez AmigoNo ratings yet
- Human Physiology-25 (DR Sachin Kapur)Document42 pagesHuman Physiology-25 (DR Sachin Kapur)AmitNo ratings yet
- Miscarriage (Abortion) : (Adhesions Inside The Uterine Cavity)Document10 pagesMiscarriage (Abortion) : (Adhesions Inside The Uterine Cavity)Rashed Shatnawi100% (1)
- Zoological Studies: An International JournalDocument142 pagesZoological Studies: An International JournalMizzannul HalimNo ratings yet
- Understanding Asexual vs Sexual ReproductionDocument5 pagesUnderstanding Asexual vs Sexual ReproductionJah Jah Dalin Lpt100% (1)
- Biology Project: PollinationDocument14 pagesBiology Project: PollinationArunkumar PNo ratings yet
- Anatomy For Beginners - Reproduction WorksheetDocument4 pagesAnatomy For Beginners - Reproduction WorksheetJames Dauray100% (1)
- Alonge, Mark - The Hymn To Zeus From Palaikastro. Religion and Tradition in Post-Minoan Crete (Greece) (PHD Thesis Stanford, UMI 2006, 275pp)Document275 pagesAlonge, Mark - The Hymn To Zeus From Palaikastro. Religion and Tradition in Post-Minoan Crete (Greece) (PHD Thesis Stanford, UMI 2006, 275pp)Marko MilosevicNo ratings yet
- Sci8 q4 Mod2 v4Document31 pagesSci8 q4 Mod2 v4Jay-ar RiosNo ratings yet
- Gen Bio IIDocument3 pagesGen Bio IIBelle MemoraBilya100% (1)
- 61 Irregular Verbs Pink and PurpleDocument18 pages61 Irregular Verbs Pink and PurpleRebeca HR SKNo ratings yet
- Alexander 2022 Deceptively Aligned MesaoptimizersDocument92 pagesAlexander 2022 Deceptively Aligned MesaoptimizersStamnumNo ratings yet
- Grade 8 Comparing Mitosis and Meiosis ModuleDocument13 pagesGrade 8 Comparing Mitosis and Meiosis ModuleLanny Padaoan LariozaNo ratings yet
- The Occult Forces of Sexual UnionDocument6 pagesThe Occult Forces of Sexual Unionimmortality2045_841960% (5)
- ReviewerDocument2 pagesReviewerFrank Jomari MurilloNo ratings yet
- Chapter 3: Selection and SpeciationDocument19 pagesChapter 3: Selection and SpeciationSANGGEETHA SRI A/P V.KUMAR MoeNo ratings yet
- Shukra DhatuDocument29 pagesShukra Dhatushilpa yermeNo ratings yet
- Essential parts of flowersDocument6 pagesEssential parts of flowersCASSANDRA REIGN CONSTANTINONo ratings yet
- Question Paper Unit A161 02 Modules b1 b2 b3 Higher TierDocument16 pagesQuestion Paper Unit A161 02 Modules b1 b2 b3 Higher TierAnonymous QFkCS837bNo ratings yet
- 7040 02 Que 20100511Document28 pages7040 02 Que 20100511Mohammad Mohasin SarderNo ratings yet
- Characteristics of Living Things Guided NotesDocument3 pagesCharacteristics of Living Things Guided NotesErica HartNo ratings yet