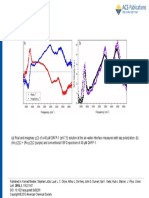
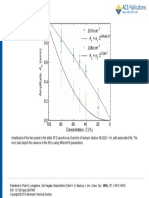
You might also like
- Butler 2018Document9 pagesButler 2018yNo ratings yet
- 10 1021-Acs Jpclett 5b00281figure3Document1 page10 1021-Acs Jpclett 5b00281figure3yNo ratings yet
- Book 2Document1 pageBook 2yNo ratings yet
- 10 1126@science Aat4010Document7 pages10 1126@science Aat4010yNo ratings yet
- 10 1021-Acs Jpclett 5b00281figure2Document1 page10 1021-Acs Jpclett 5b00281figure2yNo ratings yet
- Amber16 PDFDocument923 pagesAmber16 PDFyNo ratings yet
- What You Need To Know About Python (Ebook) - Pierluigi RitiDocument66 pagesWhat You Need To Know About Python (Ebook) - Pierluigi RitiSelf-Developer75% (4)
- Mechanisms of Glycosyl Transferases and HydrolasesDocument13 pagesMechanisms of Glycosyl Transferases and HydrolasesyNo ratings yet
- Glycosidase MechanismsDocument8 pagesGlycosidase MechanismsyNo ratings yet
- 10 1021-Jacs 5b07845figure5Document1 page10 1021-Jacs 5b07845figure5yNo ratings yet
- Qgui ManualDocument39 pagesQgui ManualyNo ratings yet
- File 1195Document86 pagesFile 1195yNo ratings yet
- 10 1021-Jacs 5b07845figure3Document1 page10 1021-Jacs 5b07845figure3yNo ratings yet
- 10 1021-Acs Jpclett 5b00281figure4Document1 page10 1021-Acs Jpclett 5b00281figure4yNo ratings yet
- 10 1021-Jacs 5b07845figure5Document1 page10 1021-Jacs 5b07845figure5yNo ratings yet
- READMEDocument1 pageREADMEyNo ratings yet
- LieDocument7 pagesLieyNo ratings yet
- 2pt Gromacs DocumentationDocument12 pages2pt Gromacs DocumentationyNo ratings yet
- Scoring of Nine Ligands in P450camDocument5 pagesScoring of Nine Ligands in P450camyNo ratings yet
- Ja5b03945 Si 001Document37 pagesJa5b03945 Si 001yNo ratings yet
- Ultrasound Velocity in WaterDocument3 pagesUltrasound Velocity in WaterAndrea EspinosaNo ratings yet
- Free Energy Perturbation AnalysisDocument12 pagesFree Energy Perturbation AnalysisyNo ratings yet
- C+Mathematica Comp PhysDocument299 pagesC+Mathematica Comp PhysyNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Confirmation Form: Pillar Regional Conference (NCR)Document1 pageConfirmation Form: Pillar Regional Conference (NCR)Llano Multi-Purpose CooperativeNo ratings yet
- 9-Lesson 5 Direct and Indirect SpeechDocument8 pages9-Lesson 5 Direct and Indirect Speechlaiwelyn100% (4)
- SO CF, Internal Fault Map Class 1A SO CF, Internal Fault Map Class 2A SO CF, External Condition Map Class 1BDocument15 pagesSO CF, Internal Fault Map Class 1A SO CF, Internal Fault Map Class 2A SO CF, External Condition Map Class 1BATMMOBILISNo ratings yet
- Syllabus Agri 2015Document248 pagesSyllabus Agri 2015parvathivignesh279No ratings yet
- E Requisition SystemDocument8 pagesE Requisition SystemWaNi AbidNo ratings yet
- The Importance of WritingDocument4 pagesThe Importance of WritingBogdan VasileNo ratings yet
- Writing A Spooky Setting DescriptionDocument4 pagesWriting A Spooky Setting DescriptionAayan AnjumNo ratings yet
- CHEE319 Tutorial 4 SolnDocument13 pagesCHEE319 Tutorial 4 SolnyeshiduNo ratings yet
- IPA Assignment Analyzes New Public AdministrationDocument8 pagesIPA Assignment Analyzes New Public AdministrationKumaran ViswanathanNo ratings yet
- Managerial Performance Evaluation ProceduresDocument3 pagesManagerial Performance Evaluation Procedures1robcortesNo ratings yet
- ERC12864-12 DemoCode 4wire SPI 2Document18 pagesERC12864-12 DemoCode 4wire SPI 2DVTNo ratings yet
- DRM 25Document14 pagesDRM 25Anantha RajanNo ratings yet
- Teodorico M. Collano, JR.: ENRM 223 StudentDocument5 pagesTeodorico M. Collano, JR.: ENRM 223 StudentJepoyCollanoNo ratings yet
- The Ethological Study of Glossifungites Ichnofacies in The Modern & Miocene Mahakam Delta, IndonesiaDocument4 pagesThe Ethological Study of Glossifungites Ichnofacies in The Modern & Miocene Mahakam Delta, IndonesiaEry Arifullah100% (1)
- Investigation of Twilight Using Sky Quality Meter For Isha' Prayer TimeDocument1 pageInvestigation of Twilight Using Sky Quality Meter For Isha' Prayer Timeresurgam52No ratings yet
- Running Head:: Describe The Uses of Waiting Line AnalysesDocument6 pagesRunning Head:: Describe The Uses of Waiting Line AnalysesHenry AnubiNo ratings yet
- 230kv Cable Sizing CalculationsDocument6 pages230kv Cable Sizing Calculationsrajinipre-1No ratings yet
- 5 Tests of Significance SeemaDocument8 pages5 Tests of Significance SeemaFinance dmsrdeNo ratings yet
- Manager - PPIC Role DescriptionDocument2 pagesManager - PPIC Role DescriptionAmit TamhankarNo ratings yet
- Unit 5 - Assessment of One'S Teaching Practice: Universidad de ManilaDocument15 pagesUnit 5 - Assessment of One'S Teaching Practice: Universidad de ManilaDoc Joey100% (3)
- Objective QuestionsDocument19 pagesObjective QuestionsDeepak SharmaNo ratings yet
- Adverb Affirmation Negation LessonDocument4 pagesAdverb Affirmation Negation LessonMire-chan Bacon100% (1)
- Rules For Assigning Activity Points: Apj Abdul Kalam Technological UniversityDocument6 pagesRules For Assigning Activity Points: Apj Abdul Kalam Technological UniversityAnonymous KyLhn6No ratings yet
- DANA 6800-1 Parts ManualDocument4 pagesDANA 6800-1 Parts ManualDude manNo ratings yet
- Reflecting on UPHSD's Mission, Vision, and Core ValuesDocument3 pagesReflecting on UPHSD's Mission, Vision, and Core ValuesBia N Cz100% (1)
- Dball-Gm5 en Ig Cp20110328aDocument18 pagesDball-Gm5 en Ig Cp20110328aMichael MartinezNo ratings yet
- Alside Brochure - Zen Windows The TriangleDocument13 pagesAlside Brochure - Zen Windows The TriangleZenWindowsTheTriangleNo ratings yet
- Rúbrica de composición escritaDocument2 pagesRúbrica de composición escritafrancisco alonsoNo ratings yet
- Lab 1 Handout Electrical MachinesDocument23 pagesLab 1 Handout Electrical Machinesvishalsharma08No ratings yet
- Ninja's Guide To OnenoteDocument13 pagesNinja's Guide To Onenotesunil100% (1)