You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Alternative Telomere RepaiirDocument16 pagesAlternative Telomere RepaiirMaithreyanNandhakumarNo ratings yet
- The Present and Future Role of Microfluidics PDFDocument9 pagesThe Present and Future Role of Microfluidics PDFMaithreyanNandhakumarNo ratings yet
- Mechanotransduction PDFDocument8 pagesMechanotransduction PDFMaithreyanNandhakumarNo ratings yet
- 4A Microfluidic 3D in Vitro Model For Specificity of Breast CancerDocument18 pages4A Microfluidic 3D in Vitro Model For Specificity of Breast CancerMaithreyanNandhakumarNo ratings yet
- Aikat1979 PDFDocument5 pagesAikat1979 PDFMaithreyanNandhakumarNo ratings yet
- Current Concept Same Bias IsDocument10 pagesCurrent Concept Same Bias IsMaithreyanNandhakumarNo ratings yet
- ThreeChannel PDFDocument8 pagesThreeChannel PDFMaithreyanNandhakumarNo ratings yet
- Extraintestinal Amebiasis': Scess Was PresentDocument3 pagesExtraintestinal Amebiasis': Scess Was PresentMaithreyanNandhakumarNo ratings yet
- 5microfluidic Assay For Simultaneous Culture of Multiple Cell Types On Surfaces or Within HydrogelsDocument29 pages5microfluidic Assay For Simultaneous Culture of Multiple Cell Types On Surfaces or Within HydrogelsMaithreyanNandhakumarNo ratings yet
- 3microfluidic Co-Culture System For Cancer Migratory Analysis and Anti-Metastatic Drugs ScreeningDocument11 pages3microfluidic Co-Culture System For Cancer Migratory Analysis and Anti-Metastatic Drugs ScreeningMaithreyanNandhakumarNo ratings yet
- 2investigating Metastasis Using in Vitro Platforms - Madame Curie Bioscience Database - NCBI BookshelfDocument25 pages2investigating Metastasis Using in Vitro Platforms - Madame Curie Bioscience Database - NCBI BookshelfMaithreyanNandhakumarNo ratings yet
- Fate of Entamoeba Histolytica During Establishment of Amoebic Liver Abscess Analyzed by Quantitative Radioimaging and HistologyDocument8 pagesFate of Entamoeba Histolytica During Establishment of Amoebic Liver Abscess Analyzed by Quantitative Radioimaging and HistologyMaithreyanNandhakumarNo ratings yet
- ReferencesDocument4 pagesReferencesMaithreyanNandhakumarNo ratings yet
- Antifreeze Proteins Enhance Survival of Cells in Cryopreservation - Substituting DMSO With RmAFP#1 in Cryopreservation of Cells 1.2Document93 pagesAntifreeze Proteins Enhance Survival of Cells in Cryopreservation - Substituting DMSO With RmAFP#1 in Cryopreservation of Cells 1.2MaithreyanNandhakumarNo ratings yet
- Introduction To Flow Cytometry - A Learning GuideDocument54 pagesIntroduction To Flow Cytometry - A Learning GuideFrank FontaineNo ratings yet
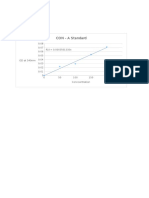
- CON - A Standard: OD at 540nmDocument10 pagesCON - A Standard: OD at 540nmMaithreyanNandhakumarNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Biol 460 Case StudyDocument11 pagesBiol 460 Case Studyapi-353888678No ratings yet
- Coccidia 10 - 11Document40 pagesCoccidia 10 - 11microperadeniya100% (1)
- PCR 5 Lab SetupDocument22 pagesPCR 5 Lab Setupptz1965No ratings yet
- Dengue FeverDocument8 pagesDengue FevervikasmaeNo ratings yet
- EmbeddingDocument6 pagesEmbeddingVikash KumarNo ratings yet
- Ace Microbiology with Active LearningDocument39 pagesAce Microbiology with Active Learning4negero33% (3)
- Nephrology MCQSDocument16 pagesNephrology MCQSThana AlAnsari100% (5)
- Nocturnal EnuresisDocument38 pagesNocturnal EnuresisayshuyNo ratings yet
- Understanding Autoimmune Disease PDFDocument18 pagesUnderstanding Autoimmune Disease PDFLiz TaylorNo ratings yet
- Intermediate MSUD: Maple Syrup Urine Disease (MSUD)Document1 pageIntermediate MSUD: Maple Syrup Urine Disease (MSUD)Awais_123No ratings yet
- Nucleic Acid and Miscellaneous Drug Targets MidtermsDocument7 pagesNucleic Acid and Miscellaneous Drug Targets MidtermsCarmela Nadine Grace CuaresmaNo ratings yet
- Nervous System WebquestDocument6 pagesNervous System Webquestapi-294104473No ratings yet
- Long Non-Coding RNAs in CancerDocument383 pagesLong Non-Coding RNAs in Cancerسہاجہدةه لہلہهNo ratings yet
- Muscular SystemDocument20 pagesMuscular SystemAltaf Hussain KhanNo ratings yet
- 15 The Neuropsychology of Dreams: A Clinico-Anatomical StudyDocument4 pages15 The Neuropsychology of Dreams: A Clinico-Anatomical StudyMaximiliano Portillo0% (1)
- Functions of Blood and ComponentsDocument15 pagesFunctions of Blood and ComponentsayuNo ratings yet
- Molecular Detection of Extended Spectrum B-Lactamases, MetalloDocument7 pagesMolecular Detection of Extended Spectrum B-Lactamases, MetalloValentina RondonNo ratings yet
- A Network Analysis of COVID-19 mRNA Vaccine Patents: ModernaDocument3 pagesA Network Analysis of COVID-19 mRNA Vaccine Patents: ModernaMatthieu BecquartNo ratings yet
- Histology: GIST Short Note by by S.Wichien (SNG KKU)Document3 pagesHistology: GIST Short Note by by S.Wichien (SNG KKU)URo KkuNo ratings yet
- ( ( ( ( ( ( ( ( ( ( ( ( ( ( ( - Clinical Trial - (Publication Type) or - Clinical Trials - PubMed - NCBI45Document23 pages( ( ( ( ( ( ( ( ( ( ( ( ( ( ( - Clinical Trial - (Publication Type) or - Clinical Trials - PubMed - NCBI45Túlio Martins OliveiraNo ratings yet
- Water BugggssDocument20 pagesWater BugggssShayna Evangelista Del ValleNo ratings yet
- Syllabus-Neuroscience 10 Neuroscience in 21st Century-2012FallDocument4 pagesSyllabus-Neuroscience 10 Neuroscience in 21st Century-2012FallJerikko Timm AgatepNo ratings yet
- Ers 2015Document358 pagesErs 2015Dejan ŽujovićNo ratings yet
- Case Conceptualization BowenDocument2 pagesCase Conceptualization BowenJovencio Marquez0% (1)
- Kumar Et Al 2008Document16 pagesKumar Et Al 2008Ruth CastilloNo ratings yet
- Case History TakingDocument30 pagesCase History Takingvenu238No ratings yet
- Nutrients 08 00068Document29 pagesNutrients 08 00068Goran TomićNo ratings yet
- About The Measure Domain MeasureDocument3 pagesAbout The Measure Domain MeasureMaríaA.SerranoNo ratings yet
- Salivary Gland - Part3Document27 pagesSalivary Gland - Part3Prince AhmedNo ratings yet
- 1.1 Fields and Careers in BiologyDocument5 pages1.1 Fields and Careers in BiologyFHATIN AMIRA BINTI MUSA MoeNo ratings yet