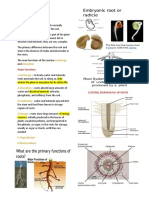
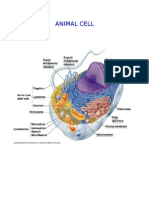
You might also like
- Chapter 35Document9 pagesChapter 35Jyothis James100% (2)
- Plant Structure and FunctionDocument14 pagesPlant Structure and FunctionPatrickNantesSalvani100% (1)
- Chapter 20 RQDocument3 pagesChapter 20 RQMia MistypuffNo ratings yet
- Lab 3 Report Plant ANATOMYDocument9 pagesLab 3 Report Plant ANATOMYEizuwanEizuddin0% (1)
- Lesson PlanDocument4 pagesLesson Planapi-289848178No ratings yet
- Stems Lesson 1: Stem FunctionsDocument11 pagesStems Lesson 1: Stem Functionskesyah rubidoNo ratings yet
- STONE (Roots&Shoots)Document3 pagesSTONE (Roots&Shoots)Allen KimaniNo ratings yet
- Botn 21n - Exercise 12 - Stems and Secondary GrowthDocument17 pagesBotn 21n - Exercise 12 - Stems and Secondary GrowthABIEGAIL LAMBERTNo ratings yet
- Last Huraahhhhh.. Go Gurl 8 - BusayDocument20 pagesLast Huraahhhhh.. Go Gurl 8 - BusayMarie Angelie AcapNo ratings yet
- ROOTSDocument10 pagesROOTSKaren ParkNo ratings yet
- RootDocument40 pagesRootAstha MishraNo ratings yet
- Anatomy of Flowering Plants - Shobhit NirwanDocument15 pagesAnatomy of Flowering Plants - Shobhit NirwanPrakhar SrivastavaNo ratings yet
- Xi-A of Atomic Energy Central School HasDocument27 pagesXi-A of Atomic Energy Central School Hasashok parmarNo ratings yet
- Plant Organ Systems Learning ObjectivesDocument12 pagesPlant Organ Systems Learning ObjectivesJoebini Aleta RobleteNo ratings yet
- Plant PhysiologyDocument11 pagesPlant PhysiologyRonniedz LicotNo ratings yet
- Comparision of The Vascular Tissue in Monocots andDocument12 pagesComparision of The Vascular Tissue in Monocots andnicoleNo ratings yet
- Villanueva - Bot01 Lab Act #4Document14 pagesVillanueva - Bot01 Lab Act #4Kath ErineNo ratings yet
- Jaringan Tumbuhan Dalam Bahasa InggrisDocument5 pagesJaringan Tumbuhan Dalam Bahasa InggrisRijal Algifari100% (1)
- Anatomy of Flowering PlantsDocument14 pagesAnatomy of Flowering PlantsRaQtrNo ratings yet
- Plant Physiology LAB REPORT XDocument5 pagesPlant Physiology LAB REPORT XBoyd benson kayomboNo ratings yet
- Mti 820003080 200323150639Document21 pagesMti 820003080 200323150639Najira AiniNo ratings yet
- Anatomy.. Paper ADocument6 pagesAnatomy.. Paper ASoha Ch100% (1)
- Anatomy of Flowering Plants NotesDocument9 pagesAnatomy of Flowering Plants NotesVansh SinghNo ratings yet
- Plant TissuesDocument6 pagesPlant TissuesJEVRIX RAMOSNo ratings yet
- Plant Histology and AnatomyDocument39 pagesPlant Histology and AnatomyN Madhusudhan50% (2)
- Anatomy of StemDocument15 pagesAnatomy of StemMonica LumbantobingNo ratings yet
- Primary Dicot Stem and Root in SunflowerDocument10 pagesPrimary Dicot Stem and Root in SunflowerPushkar kumarNo ratings yet
- Anatomy of Flowering PlantsDocument10 pagesAnatomy of Flowering Plantsl8627352No ratings yet
- Strucrtures of Plant PartsDocument95 pagesStrucrtures of Plant Partsfatima100% (1)
- Chapter 6.anatonomy of Flowering PlantsDocument12 pagesChapter 6.anatonomy of Flowering PlantsKeerthana IX-C [370]No ratings yet
- Anatomy of Flowering Plants-Class 11Document7 pagesAnatomy of Flowering Plants-Class 11ICSE BOARDNo ratings yet
- 11 Biology Notes Ch06 Anatomy of Flowering PlantsDocument8 pages11 Biology Notes Ch06 Anatomy of Flowering PlantsPriyanshu BhadanaNo ratings yet
- Plant Tissue SystemsDocument2 pagesPlant Tissue SystemsJohn0% (1)
- 6.anatomy of Flowering Plants-FDocument28 pages6.anatomy of Flowering Plants-Fshrutianand8915No ratings yet
- Chapter 6 Anatomy of Flowering Plant CompressedDocument7 pagesChapter 6 Anatomy of Flowering Plant Compressed65vxdx9ddgNo ratings yet
- Chapter - 6 Anatomy of Flowering PlantsDocument6 pagesChapter - 6 Anatomy of Flowering PlantsPriyansh PatelNo ratings yet
- Plant AnatomyDocument17 pagesPlant AnatomyPradyuman Choubey100% (1)
- ROOTS ReviewerDocument5 pagesROOTS ReviewerMANALANG JULIA ALEXISNo ratings yet
- Chapter 6 Points To RememberDocument7 pagesChapter 6 Points To RememberNANDA KISHORE S X A AD : 1 0 6 4 4No ratings yet
- The Plant BodyDocument2 pagesThe Plant BodyIJ SeducoNo ratings yet
- Root Meristem, Root Cap, and RootDocument23 pagesRoot Meristem, Root Cap, and RootJulian ManongdoNo ratings yet
- Plant Anatomy Hand Written NoteDocument95 pagesPlant Anatomy Hand Written NoteDev raj regmiNo ratings yet
- Reinforcement Plant Growth: Beaconhouse School System Unit: Class: VIIDocument8 pagesReinforcement Plant Growth: Beaconhouse School System Unit: Class: VIIAleena Zahra Naqvi/TCHR/BMBNo ratings yet
- AAPostLab The Plant BodyDocument87 pagesAAPostLab The Plant BodyAlla KanaNo ratings yet
- LeafDocument50 pagesLeafJawher AliNo ratings yet
- TissuesDocument16 pagesTissuessheruhi35No ratings yet
- Plant Tissues and The Multicellular Plant BodyDocument4 pagesPlant Tissues and The Multicellular Plant BodyAnonymous HXLczq3100% (1)
- Tissues System of The Plant BodyDocument47 pagesTissues System of The Plant BodyJana Natassia SilvaNo ratings yet
- CBSE Class 11 Biology Chapter 6 Anatomy of Flowering Plants Revision NotesDocument72 pagesCBSE Class 11 Biology Chapter 6 Anatomy of Flowering Plants Revision Noteschakrabortytrisha.2006No ratings yet
- Structures and Functions in Plants: Roots, Stems, and LeavesDocument32 pagesStructures and Functions in Plants: Roots, Stems, and LeavesDanielle AnonuevoNo ratings yet
- Plant Form and FunctionDocument32 pagesPlant Form and FunctionPaula SanicoNo ratings yet
- Plant Anatomy Class 12Document63 pagesPlant Anatomy Class 12Ranjeet Pandey100% (1)
- Meristematic TissuesDocument4 pagesMeristematic TissuesNur Syiffa RosmanNo ratings yet
- Structures and Functions in PlantsDocument32 pagesStructures and Functions in PlantsAlvin PaboresNo ratings yet
- Anatomy of Flowering PlantsDocument16 pagesAnatomy of Flowering Plantsmothi26_pcmNo ratings yet
- Botany MidtermsDocument16 pagesBotany MidtermsAnisa AzisNo ratings yet
- GS BIO 206. Functional Organization in PlantsDocument9 pagesGS BIO 206. Functional Organization in PlantsChary Johanne MenesesNo ratings yet
- The Plant Body: Tissues and MeristemDocument11 pagesThe Plant Body: Tissues and MeristemClarissa CuñaNo ratings yet
- Selaginella (Club Moss or Spike Moss) : LycopsidaDocument29 pagesSelaginella (Club Moss or Spike Moss) : LycopsidaAkshay KannanNo ratings yet
- Plant Anatomy and PhysiologyDocument33 pagesPlant Anatomy and Physiologymneilg100% (1)
- Panel 21 01HigherPlantsDocument2 pagesPanel 21 01HigherPlantsJinyuanLowNo ratings yet
- Anatomy of Flowering PlantsDocument40 pagesAnatomy of Flowering PlantsChaitali DeshmukhNo ratings yet
- How Plants Grow - With Information on the Biology of Plant Cells, Roots, Leaves and FlowersFrom EverandHow Plants Grow - With Information on the Biology of Plant Cells, Roots, Leaves and FlowersNo ratings yet
- Pavlou 2007 Pupils 39282Document26 pagesPavlou 2007 Pupils 39282Karl Chan UyNo ratings yet
- Analysis of Science Process Skills On Science LearDocument9 pagesAnalysis of Science Process Skills On Science LearKarl Chan UyNo ratings yet
- Lesson 5 Week 5Document7 pagesLesson 5 Week 5Karl Chan UyNo ratings yet
- Lesson 2 Week 2Document9 pagesLesson 2 Week 2Karl Chan UyNo ratings yet
- Lesson 4 Week 4Document4 pagesLesson 4 Week 4Karl Chan UyNo ratings yet
- Students' Science Process Skill and Analytical Thinking Ability in Chemistry LearningDocument6 pagesStudents' Science Process Skill and Analytical Thinking Ability in Chemistry LearningKarl Chan UyNo ratings yet
- Research InstrumentDocument3 pagesResearch InstrumentKarl Chan UyNo ratings yet
- Lesson 1 Week 1Document7 pagesLesson 1 Week 1Karl Chan UyNo ratings yet
- 1st InfographicDocument4 pages1st InfographicKarl Chan UyNo ratings yet
- 04 LactoseDocument1 page04 LactoseKarl Chan UyNo ratings yet
- Gay Lussac'S Law Robert Boyle: Pressure Cooker Soft DrinksDocument1 pageGay Lussac'S Law Robert Boyle: Pressure Cooker Soft DrinksKarl Chan UyNo ratings yet
- Activity On Earth and Environmental Sciences Willard UyDocument3 pagesActivity On Earth and Environmental Sciences Willard UyKarl Chan UyNo ratings yet
- 1st InfographicDocument4 pages1st InfographicKarl Chan UyNo ratings yet
- The Problem of Youth Offenders FactsDocument5 pagesThe Problem of Youth Offenders FactsKarl Chan UyNo ratings yet
- Case Digest Ucpb Et - Al V E. GanzonDocument4 pagesCase Digest Ucpb Et - Al V E. GanzonKarl Chan UyNo ratings yet
- Robin Wood Term PaperDocument9 pagesRobin Wood Term PaperKarl Chan UyNo ratings yet
- Pines City CollegesDocument2 pagesPines City CollegesKarl Chan UyNo ratings yet
- Atchie PNPDocument2 pagesAtchie PNPKarl Chan UyNo ratings yet
- EntomologyDocument6 pagesEntomologyKarl Chan UyNo ratings yet
- Robinwood Medyo Final NaDocument7 pagesRobinwood Medyo Final NaKarl Chan UyNo ratings yet
- Atchie CASESDocument87 pagesAtchie CASESKarl Chan UyNo ratings yet
- Pale Case Digest 1Document8 pagesPale Case Digest 1Karl Chan UyNo ratings yet
- Nomenclature: Hibiscus Is A Genus of Flowering Plants in The Mallow Family, Malvaceae. The Genus IsDocument4 pagesNomenclature: Hibiscus Is A Genus of Flowering Plants in The Mallow Family, Malvaceae. The Genus IsKarl Chan UyNo ratings yet
- Moises Laya JADocument5 pagesMoises Laya JAKarl Chan UyNo ratings yet
- OCA Circular No. 110 2014Document1 pageOCA Circular No. 110 2014Robert Labe JrNo ratings yet
- OCA Circular No. 110 2014Document1 pageOCA Circular No. 110 2014Robert Labe JrNo ratings yet
- GRP-LSE-008A-Anti-bribery Policy (Long Form) - CD Changes 26 Oct 16Document20 pagesGRP-LSE-008A-Anti-bribery Policy (Long Form) - CD Changes 26 Oct 16Karl Chan UyNo ratings yet
- Atchie CASESDocument87 pagesAtchie CASESKarl Chan UyNo ratings yet
- Intro To Economics With Land Reform and TaxationDocument2 pagesIntro To Economics With Land Reform and TaxationKarl Chan UyNo ratings yet
- The Problem of Youth Offenders FactsDocument5 pagesThe Problem of Youth Offenders FactsKarl Chan UyNo ratings yet
- Large Leaves Have A Large Surface Area, Which Would Allow Them To AbsorbDocument5 pagesLarge Leaves Have A Large Surface Area, Which Would Allow Them To AbsorbKeziah VenturaNo ratings yet
- Grade 10 Biology Week 1 Lesson 2 Worksheet 2 and SolutionsDocument3 pagesGrade 10 Biology Week 1 Lesson 2 Worksheet 2 and SolutionsJamie LeNo ratings yet
- Develomental Plant AnatomyDocument32 pagesDevelomental Plant AnatomyFito OrtegaNo ratings yet
- Plant Tissues QuestionsDocument2 pagesPlant Tissues QuestionsAnri van ZylNo ratings yet
- Activity 9 Secondary Growth of Roots and StemsDocument10 pagesActivity 9 Secondary Growth of Roots and StemsGenghis SupnetNo ratings yet
- Papermaking Raw Materia & Their Characteristics IIDocument115 pagesPapermaking Raw Materia & Their Characteristics IIvivekbhuchemNo ratings yet
- Lab Rep 4Document27 pagesLab Rep 4Reign RosadaNo ratings yet
- Plant Anatomy and PhysiologyDocument33 pagesPlant Anatomy and Physiologymneilg100% (1)
- Jawapan: 1.1 Organisasi Tisu Tumbuhan 1 2Document29 pagesJawapan: 1.1 Organisasi Tisu Tumbuhan 1 2ThimNo ratings yet
- Animal CellDocument5 pagesAnimal CellNan Fen Jin Sabolarce100% (1)
- APEX The Organ SystemsDocument171 pagesAPEX The Organ Systemsjt100% (2)
- Anatomy of Flowering Plants Full Chapter NotesDocument69 pagesAnatomy of Flowering Plants Full Chapter NotesmonikaNo ratings yet
- Senecio Grandidentatus Ledeb. (Asteraceae) : Anatomical Aspects of The Stem and Leaf ofDocument7 pagesSenecio Grandidentatus Ledeb. (Asteraceae) : Anatomical Aspects of The Stem and Leaf ofBlinda Raluca MariaNo ratings yet
- SONI - Fundamentals of Botany - Vol 2 (2011, MC GRAW HILL INDIA) PDFDocument398 pagesSONI - Fundamentals of Botany - Vol 2 (2011, MC GRAW HILL INDIA) PDFPrince Quokka Rai100% (5)
- Anatomy & Plant PhysiologyDocument27 pagesAnatomy & Plant Physiologyshree devNo ratings yet
- Anatomy of Flowering PlantsDocument16 pagesAnatomy of Flowering Plantsmothi26_pcmNo ratings yet
- Plant Histology and AnatomyDocument39 pagesPlant Histology and AnatomyN Madhusudhan50% (2)
- BIOL1012 Module 5 - Part 1 Morphology, Internal Structure and Growth of PlantsDocument79 pagesBIOL1012 Module 5 - Part 1 Morphology, Internal Structure and Growth of PlantsKayla CoxNo ratings yet
- Early Flowers and Angiosperm EvolutionDocument597 pagesEarly Flowers and Angiosperm EvolutionwccardosoNo ratings yet
- FC101 Act.5 Root Morphology and AnatomyDocument8 pagesFC101 Act.5 Root Morphology and AnatomyCristine egagamaoNo ratings yet
- Physiology of Pruning Fruit Trees VCE 422-025Document8 pagesPhysiology of Pruning Fruit Trees VCE 422-025Anonymous aSefOrQ9I100% (1)
- Bot 307 Comparative Plant Anatomy 1Document62 pagesBot 307 Comparative Plant Anatomy 1Kennedy Samuel BanjaNo ratings yet
- Plant AnatomyDocument22 pagesPlant AnatomyYogesh Bagad100% (1)
- Botany Compare and ContrastDocument6 pagesBotany Compare and ContrastMa. Ellah Patricia M. GutierrezNo ratings yet
- Dokumen - Tips Lab 3 Report Plant AnatomyDocument11 pagesDokumen - Tips Lab 3 Report Plant AnatomyAin SufizaNo ratings yet
- Practical Key-2023Document5 pagesPractical Key-2023aizah thameemNo ratings yet