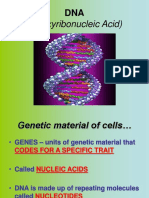
You might also like
- Central Dogma of Molecular BiologyDocument9 pagesCentral Dogma of Molecular BiologyXavier Langgaman100% (1)
- Genetic Information: - Central Dogma - Genetic CodesDocument34 pagesGenetic Information: - Central Dogma - Genetic Codesakhikabir100% (1)
- Central DogmaDocument50 pagesCentral DogmaIvilyn ManguladNo ratings yet
- FCB 7-GeneticsDocument15 pagesFCB 7-GeneticsYuri PaderesNo ratings yet
- Transport in PlantsDocument21 pagesTransport in PlantsMothi KarunaNo ratings yet
- Zoology Lab Manual Hard CoverDocument103 pagesZoology Lab Manual Hard CoverMona Saad AliNo ratings yet
- Molecular Cell Biology 1 Basics of Molecular BiologyDocument80 pagesMolecular Cell Biology 1 Basics of Molecular BiologyIana Mira100% (2)
- DNA Structure and Function - First Lecture (4822)Document17 pagesDNA Structure and Function - First Lecture (4822)Ashley Ramos LópezNo ratings yet
- Replication, Transcription, Translation and Its Regulation: By, University of Agricultural Sciences, DharwadDocument42 pagesReplication, Transcription, Translation and Its Regulation: By, University of Agricultural Sciences, DharwadTabada NickyNo ratings yet
- Protein Synthesis SummaryDocument2 pagesProtein Synthesis SummaryTrinitee Wesner100% (1)
- Central Dogma of Molecular BiologyDocument7 pagesCentral Dogma of Molecular BiologyRenee Louise CasullaNo ratings yet
- Bio 106 Lecture 5 Chemical Basis of Heredit1Document16 pagesBio 106 Lecture 5 Chemical Basis of Heredit1Evan Jane JumamilNo ratings yet
- Central Dogma of Molecular BiologyDocument48 pagesCentral Dogma of Molecular BiologyLJ Jamin CentinoNo ratings yet
- Recombinant Dna TechnologyDocument3 pagesRecombinant Dna TechnologyAeron Jade ParenaNo ratings yet
- DNA ReplicationDocument5 pagesDNA Replicationvijayalakshmiraman100% (1)
- DNA and PROTEIN SYNTHESISDocument60 pagesDNA and PROTEIN SYNTHESISathirah100% (1)
- Protein Synthesis: Class Notes NotesDocument2 pagesProtein Synthesis: Class Notes NotesDale HardingNo ratings yet
- Steps of DNA ReplicationDocument5 pagesSteps of DNA Replicationatul mishra100% (1)
- Biodiversity and Classification NLDocument23 pagesBiodiversity and Classification NLruxan jean abalos100% (1)
- Syllabus in Chemical Biology III (Biomolecules) (TCB3)Document12 pagesSyllabus in Chemical Biology III (Biomolecules) (TCB3)Yuri PaderesNo ratings yet
- Cell DifferentiationDocument16 pagesCell DifferentiationHarika KeshavNo ratings yet
- Central Dogma ActivityDocument5 pagesCentral Dogma ActivitySamuel ZhangNo ratings yet
- Protein Synthesis Notes and DiagramDocument34 pagesProtein Synthesis Notes and Diagramapi-267117865100% (1)
- Lab Activity No. 12 Phylogenetic SystematicsfDocument11 pagesLab Activity No. 12 Phylogenetic SystematicsfMelanie Mamon100% (1)
- Genetic EngineeringDocument218 pagesGenetic EngineeringSrramNo ratings yet
- The Central Dogma of Molecular BiologyDocument45 pagesThe Central Dogma of Molecular BiologyEirla Kneriz Trinidad67% (3)
- Central Dogma of Molecular GeneticsDocument3 pagesCentral Dogma of Molecular GeneticsDiana RomeroNo ratings yet
- Study Guide Protein Synthesis (KEY)Document2 pagesStudy Guide Protein Synthesis (KEY)owls_1102No ratings yet
- Cell BiologyDocument16 pagesCell BiologyKlint Rodney Mark Lule100% (1)
- Recombinant DNA TechnologyDocument24 pagesRecombinant DNA TechnologySajjad AhmadNo ratings yet
- 6.LECTURE 6 - TaxonomyDocument35 pages6.LECTURE 6 - TaxonomyMuhammad Danial HasanNo ratings yet
- ChromosomesDocument39 pagesChromosomesSuresh Kannan100% (1)
- Central Dogma of Molecular BiologyDocument23 pagesCentral Dogma of Molecular BiologyEros James Jamero100% (1)
- Cellular Molecular BiologyDocument4 pagesCellular Molecular BiologyCpopNo ratings yet
- Deoxyribonucleic AcidDocument36 pagesDeoxyribonucleic AcidAlex LlanoNo ratings yet
- Basic Genetics 1Document37 pagesBasic Genetics 1Satyasundar MohantyNo ratings yet
- Chapter 2 - VirusDocument26 pagesChapter 2 - Virusshahera rosdiNo ratings yet
- Phylogenetic Tree: Tree Is A Branching Diagram or "Tree"Document6 pagesPhylogenetic Tree: Tree Is A Branching Diagram or "Tree"Juan Santiago Pineda Rodriguez PequeNo ratings yet
- Cel Mol LecDocument3 pagesCel Mol LecIzza LimNo ratings yet
- The Cell Cycle ReviewerDocument3 pagesThe Cell Cycle ReviewerJoelle Noriko G. Galang0% (1)
- Central Dogma ReviewerDocument7 pagesCentral Dogma ReviewerCarmela LipoNo ratings yet
- Miller/Tupper: Zoology 11e Instructor's Manual: Chapter SummaryDocument3 pagesMiller/Tupper: Zoology 11e Instructor's Manual: Chapter SummaryCaesar Franz RuizNo ratings yet
- PhylogenyDocument43 pagesPhylogenyMausam KumravatNo ratings yet
- Recombinant DNA TechnologyDocument26 pagesRecombinant DNA TechnologyChrystel Comendaodr100% (1)
- Phylogenetic Trees Click Learn WorksheetDocument4 pagesPhylogenetic Trees Click Learn Worksheetfcm31450% (1)
- Ch5 Cell MembranesDocument60 pagesCh5 Cell MembranesAdrian Aguilera GarciaNo ratings yet
- Chapter 10 Test BankDocument11 pagesChapter 10 Test Bankanon_747148947No ratings yet
- Introduction To Biotechnology PDFDocument39 pagesIntroduction To Biotechnology PDFFitaSucia100% (1)
- Basic Cell Types, Cell Structure and Function,& Cell ModificationsDocument28 pagesBasic Cell Types, Cell Structure and Function,& Cell ModificationsMary Ann Gonzales Abeñon100% (1)
- Photosynthesis Study GuideDocument9 pagesPhotosynthesis Study GuideJunhee KimNo ratings yet
- Bt0213 - Cell Biology Practical ManualDocument31 pagesBt0213 - Cell Biology Practical ManualVenus Divinagracia0% (2)
- Transgenic Animals: Their Benefits To Human Welfare: What Is A Transgenic Animal?Document7 pagesTransgenic Animals: Their Benefits To Human Welfare: What Is A Transgenic Animal?anon_939844203No ratings yet
- 15 The Cell Cycle-S PDFDocument6 pages15 The Cell Cycle-S PDFAml Aml0% (1)
- Cell Membrane StructureDocument17 pagesCell Membrane StructurearunNo ratings yet
- VirusDocument57 pagesVirusSophan HadieNo ratings yet
- Nucleic Acids-DNA and RNADocument12 pagesNucleic Acids-DNA and RNADingdong-didiNo ratings yet
- DNA Replication PresentationDocument28 pagesDNA Replication PresentationCelely Nitnit100% (2)
- Prodcut of Recombinat DNA TechnologyDocument9 pagesProdcut of Recombinat DNA TechnologySHOAIB NAVEEDNo ratings yet
- 2.5.) Central Dogma of Molecular Biology (BIO40)Document21 pages2.5.) Central Dogma of Molecular Biology (BIO40)Mary Ferl Jasmin LupagueNo ratings yet
- Central DogmaDocument17 pagesCentral DogmaPrakash KumarNo ratings yet
- Pumps, Compressors and Conveyors - Rev4Document179 pagesPumps, Compressors and Conveyors - Rev4Kim GojoCruzNo ratings yet
- Philippine HistoryDocument34 pagesPhilippine HistoryKim GojoCruzNo ratings yet
- Pumps, Compressors and Conveyors - KimDocument42 pagesPumps, Compressors and Conveyors - KimKim GojoCruzNo ratings yet
- HumidificationDocument29 pagesHumidificationClark Ivan Torres100% (3)
- Gas Absorption PDFDocument42 pagesGas Absorption PDFKim GojoCruz90% (30)
- Crystallization NotesDocument42 pagesCrystallization NotesKTINE0894% (16)
- Evaporation: Chemical Engineering SeriesDocument52 pagesEvaporation: Chemical Engineering Serieskmrosario91% (32)
- Drying: Chemical Engineering SeriesDocument54 pagesDrying: Chemical Engineering Serieskmrosario67% (21)
- DryingDocument12 pagesDryingKim GojoCruzNo ratings yet
- Unit 4Document2 pagesUnit 4Sweta YadavNo ratings yet
- DockerDocument35 pagesDocker2018pgicsankush10No ratings yet
- PHP Listado de EjemplosDocument137 pagesPHP Listado de Ejemploslee9120No ratings yet
- Fundamental of Transport PhenomenaDocument521 pagesFundamental of Transport Phenomenaneloliver100% (2)
- RBConcept Universal Instruction ManualDocument19 pagesRBConcept Universal Instruction Manualyan henrique primaoNo ratings yet
- B. Complete The Following Exercise With or Forms of The Indicated VerbsDocument4 pagesB. Complete The Following Exercise With or Forms of The Indicated VerbsLelyLealGonzalezNo ratings yet
- Articles About Gossip ShowsDocument5 pagesArticles About Gossip ShowssuperultimateamazingNo ratings yet
- Gothic Fiction Oliver TwistDocument3 pagesGothic Fiction Oliver TwistTaibur RahamanNo ratings yet
- MSDS Blattanex GelDocument5 pagesMSDS Blattanex GelSadhana SentosaNo ratings yet
- Indian Retail Industry: Structure, Drivers of Growth, Key ChallengesDocument15 pagesIndian Retail Industry: Structure, Drivers of Growth, Key ChallengesDhiraj YuvrajNo ratings yet
- Two Gentlemen of VeronaDocument13 pagesTwo Gentlemen of Veronavipul jainNo ratings yet
- Catibog Approval Sheet EditedDocument10 pagesCatibog Approval Sheet EditedCarla ZanteNo ratings yet
- Net 2020Document48 pagesNet 2020Krishan pandeyNo ratings yet
- Created by EDGAR Online, Inc. Dow Chemical Co /de/ Table - of - Contents Form Type: 10-K Period End: Dec 31, 2020 Date Filed: Feb 05, 2021Document286 pagesCreated by EDGAR Online, Inc. Dow Chemical Co /de/ Table - of - Contents Form Type: 10-K Period End: Dec 31, 2020 Date Filed: Feb 05, 2021Udit GuptaNo ratings yet
- Peace Corps Samoa Medical Assistant Office of The Public Service of SamoaDocument10 pagesPeace Corps Samoa Medical Assistant Office of The Public Service of SamoaAccessible Journal Media: Peace Corps DocumentsNo ratings yet
- A1 - The Canterville Ghost WorksheetsDocument8 pagesA1 - The Canterville Ghost WorksheetsТатьяна ЩукинаNo ratings yet
- Upload A Document To Access Your Download: Social Studies of HealthDocument3 pagesUpload A Document To Access Your Download: Social Studies of Health1filicupENo ratings yet
- Vistas 1-7 Class - 12 Eng - NotesDocument69 pagesVistas 1-7 Class - 12 Eng - Notesvinoth KumarNo ratings yet
- Puzzles 1Document214 pagesPuzzles 1Prince VegetaNo ratings yet
- V.S.B Engineering College, KarurDocument3 pagesV.S.B Engineering College, KarurKaviyarasuNo ratings yet
- Do or Does1.1.2Document4 pagesDo or Does1.1.2dzanardipintoNo ratings yet
- Affin Bank V Zulkifli - 2006Document15 pagesAffin Bank V Zulkifli - 2006sheika_11No ratings yet
- World Price List 2014: Adventys Induction Counter TopsDocument4 pagesWorld Price List 2014: Adventys Induction Counter TopsdiogocorollaNo ratings yet
- Apforest Act 1967Document28 pagesApforest Act 1967Dgk RajuNo ratings yet
- MYPNA SE G11 U1 WebDocument136 pagesMYPNA SE G11 U1 WebKokiesuga12 TaeNo ratings yet
- ICFR Presentation - Ernst and YoungDocument40 pagesICFR Presentation - Ernst and YoungUTIE ELISA RAMADHANI67% (3)
- BCMSN30SG Vol.2 PDFDocument394 pagesBCMSN30SG Vol.2 PDFShemariyahNo ratings yet
- Physics Tadka InstituteDocument15 pagesPhysics Tadka InstituteTathagata BhattacharjyaNo ratings yet
- 7C Environment Test 2004Document2 pages7C Environment Test 2004api-3698146No ratings yet
- Diec Russias Demographic Policy After 2000 2022Document29 pagesDiec Russias Demographic Policy After 2000 2022dawdowskuNo ratings yet