You might also like
- C5RA01911GDocument55 pagesC5RA01911GAndrew LondonNo ratings yet
- DPPH Radical Scavenging AssayDocument20 pagesDPPH Radical Scavenging AssayBeatriz FriasNo ratings yet
- Antioxidants and Their Charecterization: September 2011Document4 pagesAntioxidants and Their Charecterization: September 2011SofhiaZahraNo ratings yet
- 6 Vol. 6 Issue 2 Feb 2015 IJPSR RE 1434 Paper 6Document21 pages6 Vol. 6 Issue 2 Feb 2015 IJPSR RE 1434 Paper 6Deepshikha GuptaNo ratings yet
- Methods For Determination of Antioxidant Capacity: A Review: IJPSR (2015), Vol. 6, Issue 2 (Review Article)Document21 pagesMethods For Determination of Antioxidant Capacity: A Review: IJPSR (2015), Vol. 6, Issue 2 (Review Article)Anonymous 6OPLC9UNo ratings yet
- Antioxidant Activity: An OverviewDocument11 pagesAntioxidant Activity: An Overviewsia1811No ratings yet
- Oxidative Stress, Insulin Resistance, Dyslipidemia and Type 2 Diabetes MellitusDocument26 pagesOxidative Stress, Insulin Resistance, Dyslipidemia and Type 2 Diabetes MellitusRavika Mutiara MansurNo ratings yet
- A Review On Antioxidant Methods: Dontha SunithaDocument20 pagesA Review On Antioxidant Methods: Dontha SunithaAzizah MunitaNo ratings yet
- Understanding Oxidants and Antioxidants: Classical Team With New PlayersDocument13 pagesUnderstanding Oxidants and Antioxidants: Classical Team With New PlayersjenniNo ratings yet
- Antioxidant Good PaperDocument11 pagesAntioxidant Good PaperSharon ChristantoNo ratings yet
- Foods 12 03314Document14 pagesFoods 12 03314ela.sofiaNo ratings yet
- Oxidative Stress, Aging, and Diseases: Clinical Interventions in Aging DoveDocument16 pagesOxidative Stress, Aging, and Diseases: Clinical Interventions in Aging DoveMedicina EncantadaNo ratings yet
- Natural Antioxidants Function and SourcesDocument7 pagesNatural Antioxidants Function and SourcesandreiRMSNo ratings yet
- Natural Antioxidants: Function and SourcesDocument7 pagesNatural Antioxidants: Function and SourcesBilqis NurganiyuNo ratings yet
- Oxidative Stress and AntioxidaDocument31 pagesOxidative Stress and AntioxidaJaya KumarNo ratings yet
- Antioxidan ReviewDocument9 pagesAntioxidan ReviewGustavo RuizNo ratings yet
- Antioxidant Intake and Antitumor Therapy Toward Nutritional Recommendations For Optimal Results (SaludDocument20 pagesAntioxidant Intake and Antitumor Therapy Toward Nutritional Recommendations For Optimal Results (Saludzahra tsaniaNo ratings yet
- Antioxidant Methods PDFDocument19 pagesAntioxidant Methods PDFVageeshbabu HanurNo ratings yet
- Methodology For The Determination of BioDocument11 pagesMethodology For The Determination of BioCaryl Ann C. SernadillaNo ratings yet
- A Review On: Antioxidant and Its Impact During The Bread Making ProcessDocument5 pagesA Review On: Antioxidant and Its Impact During The Bread Making ProcessDan R. A. VieiraNo ratings yet
- Review Article: Oxidative Stress Versus Antioxidants: December 2014Document13 pagesReview Article: Oxidative Stress Versus Antioxidants: December 2014Zeljko LekovicNo ratings yet
- A.A R.R.. - May. 2008 PDFDocument16 pagesA.A R.R.. - May. 2008 PDFNandagopal KNo ratings yet
- Administration of Antioxidants in Cancer-DikonversiDocument14 pagesAdministration of Antioxidants in Cancer-DikonversiTasya Aulia PNo ratings yet
- A Review of Antioxidant MethodDocument19 pagesA Review of Antioxidant Methodgiahuyle78No ratings yet
- Free Radical Scavenging Activity of Marine SpongesDocument8 pagesFree Radical Scavenging Activity of Marine SpongeshadifaNo ratings yet
- v3 308 316 PDFDocument9 pagesv3 308 316 PDFKapil SoniNo ratings yet
- Bioavailability and Antioxidant Activity of Black Chokeberry (Mechanisms of Action: A ReviewDocument19 pagesBioavailability and Antioxidant Activity of Black Chokeberry (Mechanisms of Action: A Reviewdon.meNo ratings yet
- Vivo. As Shown, There Are Several Lines of Defence. The First Defence Line IsDocument8 pagesVivo. As Shown, There Are Several Lines of Defence. The First Defence Line IsClaudia SurdilaNo ratings yet
- 8 Oxidative StressDocument13 pages8 Oxidative StressVictor ApolonioNo ratings yet
- Antioxidantes en EmbriónDocument8 pagesAntioxidantes en EmbriónCarolina PosadaNo ratings yet
- Toxicology Reports: Jonathan P. Adjimani, Prince AsareDocument8 pagesToxicology Reports: Jonathan P. Adjimani, Prince AsareBarot YashNo ratings yet
- Significance of AO Potencial of Plantas and Its Relevance To Therapeut Appl Kasote DM 2015Document10 pagesSignificance of AO Potencial of Plantas and Its Relevance To Therapeut Appl Kasote DM 2015Stephanie Aguilar TiradoNo ratings yet
- Halliwell 2008Document6 pagesHalliwell 2008Meow Vendo PermutoNo ratings yet
- Direct and Indirect Antioxidant Properties of Alpha Lipoic Acid (MECOLA)Document13 pagesDirect and Indirect Antioxidant Properties of Alpha Lipoic Acid (MECOLA)Hendri YantoNo ratings yet
- AntioxidantsDocument14 pagesAntioxidantsReena SapruNo ratings yet
- Antioxidants 09 01062 v3Document25 pagesAntioxidants 09 01062 v3melvinMAPNo ratings yet
- Preservativos Review1Document14 pagesPreservativos Review1Rubi PerezNo ratings yet
- Tocopherols As Antioxidants in Lipid Based Systems The Combination of Chemical and Physicochemical Interactions Determines Their EfficieDocument47 pagesTocopherols As Antioxidants in Lipid Based Systems The Combination of Chemical and Physicochemical Interactions Determines Their EfficieDiego MonterrozaNo ratings yet
- Free Radicals, Antioxidants and Functional Foods: Impact On Human HealthDocument10 pagesFree Radicals, Antioxidants and Functional Foods: Impact On Human HealthnurulNo ratings yet
- Antioxidants Science Technology and ApplicationsDocument61 pagesAntioxidants Science Technology and ApplicationsNguyễn Trung HiếuNo ratings yet
- Phenolic Antioxidant Capacity A Review of The Stat PDFDocument18 pagesPhenolic Antioxidant Capacity A Review of The Stat PDFSantiago TabaresNo ratings yet
- A Review On The Antioxidant and Therapeutic Potential of Bacopa MonnieriDocument12 pagesA Review On The Antioxidant and Therapeutic Potential of Bacopa MonnieriAn ThạnhNo ratings yet
- Hydrogen As A Selective Antioxidant PDFDocument12 pagesHydrogen As A Selective Antioxidant PDFIhorNo ratings yet
- Free Radicals and AntioxidantsDocument6 pagesFree Radicals and AntioxidantsAnonymous ceYk4p4No ratings yet
- Chitosan Hydrogel Nanotiter Biosensors: 指導教授:Prof. Pawel Urban 學號:108000265 學生:黃琮貽 中華民國 111 年 6 月Document27 pagesChitosan Hydrogel Nanotiter Biosensors: 指導教授:Prof. Pawel Urban 學號:108000265 學生:黃琮貽 中華民國 111 年 6 月SmileyQing ChuaNo ratings yet
- Review Article: Characterization, Preparation, and Purification of Marine Bioactive PeptidesDocument17 pagesReview Article: Characterization, Preparation, and Purification of Marine Bioactive Peptidessumaiya rahmanNo ratings yet
- Reviewof Methodsto Determine Antioxidant CapacitiesDocument21 pagesReviewof Methodsto Determine Antioxidant Capacitiesnoura farahNo ratings yet
- Materials 14 04135 v2Document54 pagesMaterials 14 04135 v2Kareem AldalyNo ratings yet
- Arouma OI, 1999Document11 pagesArouma OI, 1999Sergio mauricio sergioNo ratings yet
- Solublidad de NucleosidosDocument18 pagesSolublidad de NucleosidosLaura MartínezNo ratings yet
- Clinical Effects of Hydrogen Administration From ADocument46 pagesClinical Effects of Hydrogen Administration From AJimmy PrayogiNo ratings yet
- Antioxidants 11 02447Document29 pagesAntioxidants 11 02447Emi009No ratings yet
- Targeting Oxidative Stress in Disease: Promise and Limitations of Antioxidant TherapyDocument21 pagesTargeting Oxidative Stress in Disease: Promise and Limitations of Antioxidant TherapyMedicina EncantadaNo ratings yet
- Park 2019Document10 pagesPark 2019huyenpv91No ratings yet
- Oxidative Stress PDFDocument7 pagesOxidative Stress PDFKatarida DolluNo ratings yet
- Oxidative StressDocument13 pagesOxidative StressAnNi FitRiaNo ratings yet
- Degradation Pathway of Pharmaceutical Dosage Forms: March 2017Document26 pagesDegradation Pathway of Pharmaceutical Dosage Forms: March 2017Mihwak BlackNo ratings yet
- 2review Chikezie OxstressdiabraDocument20 pages2review Chikezie OxstressdiabraFaidil AkbarNo ratings yet
- 10 1002@jccs 201600051Document12 pages10 1002@jccs 201600051taoufik akabliNo ratings yet
- Morning HeadacheDocument7 pagesMorning HeadacheAnna DiaconuNo ratings yet
- Cognitive Impairment in Obstructive Sleep Apnea 2014 Pathologie BiologieDocument8 pagesCognitive Impairment in Obstructive Sleep Apnea 2014 Pathologie BiologieAnna DiaconuNo ratings yet
- Tulb CognitiveDocument16 pagesTulb CognitiveAnna DiaconuNo ratings yet
- Understanding The Rhythm of BreathingDocument18 pagesUnderstanding The Rhythm of BreathingAnna DiaconuNo ratings yet
- Clasificarea Tulburarilor de Somn 2Document12 pagesClasificarea Tulburarilor de Somn 2Anna DiaconuNo ratings yet
- Chapter 4 Cognitive Recovery Following Positive Airw 2011 Progress in BraiDocument18 pagesChapter 4 Cognitive Recovery Following Positive Airw 2011 Progress in BraiAnna DiaconuNo ratings yet
- The Science of Antioxidants Can Be Quite ComplexDocument3 pagesThe Science of Antioxidants Can Be Quite ComplexAnna DiaconuNo ratings yet
- The Relation Between Dose of Vitamin e And...Document6 pagesThe Relation Between Dose of Vitamin e And...Anna DiaconuNo ratings yet
- Evaluarea Practica in Pediatrie (Scan C.a.)Document67 pagesEvaluarea Practica in Pediatrie (Scan C.a.)Anna DiaconuNo ratings yet
- The Relation Between Dose of Vitamin e And...Document6 pagesThe Relation Between Dose of Vitamin e And...Anna DiaconuNo ratings yet
- The Effect of Vitamin C and E On...Document8 pagesThe Effect of Vitamin C and E On...Anna DiaconuNo ratings yet
- Gray RTSDocument10 pagesGray RTSAnna DiaconuNo ratings yet
- Free Radicals and Antioxidants in Normal PhysiologicalDocument41 pagesFree Radicals and Antioxidants in Normal PhysiologicalleosabreuNo ratings yet
- Personality Traits Vestibulo CohlearDocument10 pagesPersonality Traits Vestibulo CohlearAnna DiaconuNo ratings yet
- Peronality Teories Comparative ReviewDocument2 pagesPeronality Teories Comparative ReviewAnna DiaconuNo ratings yet
- Free Radicals and Tissue Damage Role of AntioxidantsDocument6 pagesFree Radicals and Tissue Damage Role of AntioxidantsAnna DiaconuNo ratings yet
- Peronality Teories Comparative ReviewDocument2 pagesPeronality Teories Comparative ReviewAnna DiaconuNo ratings yet
- Vitamin C and e Beneficial Effects From A Mechanistic PerspectiveDocument14 pagesVitamin C and e Beneficial Effects From A Mechanistic PerspectiveAnna DiaconuNo ratings yet
- Personality Traits AsthmaDocument10 pagesPersonality Traits AsthmaAnna DiaconuNo ratings yet
- Chest X-Ray Interpretation: Bucky Boaz, ARNP-CDocument101 pagesChest X-Ray Interpretation: Bucky Boaz, ARNP-CAnna DiaconuNo ratings yet
- Sample Size CalcDocument15 pagesSample Size CalcAnna DiaconuNo ratings yet
- ABC of Heart DiseaseDocument78 pagesABC of Heart DiseaseRolando Reyna100% (2)
- Doxycycline (As Hyclate) : Package Leaflet: Information For The User Doxycycline Pinewood 100 MG CapsulesDocument2 pagesDoxycycline (As Hyclate) : Package Leaflet: Information For The User Doxycycline Pinewood 100 MG CapsulesZulva Chairunnisa BudimanNo ratings yet
- Wa0003Document4 pagesWa0003YegaNo ratings yet
- 2nd Long Exam BlueprintDocument3 pages2nd Long Exam BlueprintRachelle MaravillaNo ratings yet
- Circadian Misalignment and HealthDocument16 pagesCircadian Misalignment and HealthGiovane PiresNo ratings yet
- Breastfeeding QuestionnaireDocument2 pagesBreastfeeding QuestionnaireDr Puteri Nur Sabrina Binti Mohd HanapiNo ratings yet
- Exercise Science Honours 2016Document22 pagesExercise Science Honours 2016asdgffdNo ratings yet
- March Case StudyDocument13 pagesMarch Case Studyapi-212894050No ratings yet
- Tromso. Norway Orders Drug-Free Treatment in PsychiatryDocument21 pagesTromso. Norway Orders Drug-Free Treatment in PsychiatryMarcelo Astorga VelosoNo ratings yet
- Daycare Fact SheetDocument1 pageDaycare Fact SheetJoana R. SilvaNo ratings yet
- Tear and Pterygium A Clinicopathological Study of Conjunctiva For Tear Film Anomaly in PterygiumDocument7 pagesTear and Pterygium A Clinicopathological Study of Conjunctiva For Tear Film Anomaly in PterygiumShofi Dhia AiniNo ratings yet
- Risk Factors For CKDDocument2 pagesRisk Factors For CKDgigolo13No ratings yet
- Nutritional Prehabilitation Program and Cardiac Surgery Outcome in PediatricsDocument15 pagesNutritional Prehabilitation Program and Cardiac Surgery Outcome in PediatricsIOSRjournalNo ratings yet
- Language and ArtsDocument17 pagesLanguage and ArtsOpium PoppyNo ratings yet
- Inles 4Document2 pagesInles 4Astrid CruzNo ratings yet
- Environmental Health: Is The Smokers Exposure To Environmental Tobacco Smoke Negligible?Document14 pagesEnvironmental Health: Is The Smokers Exposure To Environmental Tobacco Smoke Negligible?Bill HanneganNo ratings yet
- Lab 1C PAR-Q and Health History QuestionnaireDocument4 pagesLab 1C PAR-Q and Health History Questionnaireskyward23No ratings yet
- b1 ChecklistDocument2 pagesb1 ChecklistRoryNo ratings yet
- Nursing Care For BipolarDocument3 pagesNursing Care For BipolarAnthonyChapotin100% (2)
- PDFDocument732 pagesPDFMahmud Eljaarani90% (10)
- Nursing Care Plan: Student's Name: ID: Course Name: SettingDocument3 pagesNursing Care Plan: Student's Name: ID: Course Name: SettingnjoodNo ratings yet
- Clinico-Microbiological Spectrum of Hemodialysis Catheter-Related Blood Stream Infections: A Concise ReviewDocument7 pagesClinico-Microbiological Spectrum of Hemodialysis Catheter-Related Blood Stream Infections: A Concise ReviewIJAR JOURNALNo ratings yet
- Protozoa and Fungi Culturing in The LabDocument7 pagesProtozoa and Fungi Culturing in The LabrorenNo ratings yet
- Tracheal Intubation: Airway AssessmentDocument7 pagesTracheal Intubation: Airway AssessmentElle ReyesNo ratings yet
- Bee Empowerment Leslie WilsonDocument6 pagesBee Empowerment Leslie Wilsonchakravartys1327100% (1)
- Human Anatomy & Physiology: Chapter 21-1Document103 pagesHuman Anatomy & Physiology: Chapter 21-1AngelyNo ratings yet
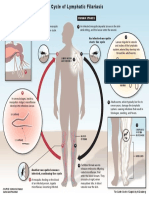
- Lymphatic Filariasis Life Cycle PDFDocument1 pageLymphatic Filariasis Life Cycle PDFlauraNo ratings yet
- Nursing Care of HydronephrosisDocument19 pagesNursing Care of Hydronephrosisvictor zhefaNo ratings yet
- CBSE Class 8 Science Sample Paper Set 5: Each Question Carriers 1 Mark: Choose The Correct Answers: (5 X 1 5 Marks)Document3 pagesCBSE Class 8 Science Sample Paper Set 5: Each Question Carriers 1 Mark: Choose The Correct Answers: (5 X 1 5 Marks)shweta kothariNo ratings yet
- Inggris 201Document5 pagesInggris 201hakimzainulNo ratings yet
- Annual Drug Data Report Vol-1 1971Document228 pagesAnnual Drug Data Report Vol-1 1971lasicablava50% (2)