You might also like
- Wound Healing SchwartzDocument37 pagesWound Healing SchwartzCarla Putri Chandra100% (1)
- School Cell Analogy PpointDocument14 pagesSchool Cell Analogy Ppointapi-265180883No ratings yet
- Chap11 LearnObjDocument4 pagesChap11 LearnObjamariliyahNo ratings yet
- Test Bank For Genetics A Conceptual Approach 4th Edition Benjamin A Pierce DownloadDocument12 pagesTest Bank For Genetics A Conceptual Approach 4th Edition Benjamin A Pierce Downloaddillonmillerjasqrpgnez100% (22)
- Cell Theory - Term PaperDocument16 pagesCell Theory - Term PaperPhyl MarylNo ratings yet
- Science 6 Chapter 1 To 12 Final Teacher Guide Final 2018 NewDocument197 pagesScience 6 Chapter 1 To 12 Final Teacher Guide Final 2018 NewAyesha SahidNo ratings yet
- Cell Adhesion MoleculesDocument90 pagesCell Adhesion MoleculesPooja Bappanna100% (1)
- Molecular BiologyDocument2 pagesMolecular BiologyJaymark HisonaNo ratings yet
- Lecture 7 BBT312 DNA Replication Part IDocument24 pagesLecture 7 BBT312 DNA Replication Part IBol BirNo ratings yet
- Module 2 Heredity - Inheritance and Variation Central DogmaDocument36 pagesModule 2 Heredity - Inheritance and Variation Central Dogmaseph20lupozNo ratings yet
- Dna Replication - Dna RepairDocument27 pagesDna Replication - Dna Repairnokate konkoorNo ratings yet
- Central Dogma of BiologyDocument6 pagesCentral Dogma of BiologyJhun Lerry TayanNo ratings yet
- Dna & RnaDocument79 pagesDna & RnaShimmering MoonNo ratings yet
- European Journal of Biochemistry - December 1990 - TH MMES - Eukaryotic DNA Replication PDFDocument14 pagesEuropean Journal of Biochemistry - December 1990 - TH MMES - Eukaryotic DNA Replication PDFFarhadullah KhanNo ratings yet
- Lesson 5 CYTO DNA ReplicationDocument4 pagesLesson 5 CYTO DNA ReplicationTherese TimbalNo ratings yet
- S Phase and DNA ReplicationDocument15 pagesS Phase and DNA Replicationdiego angelesNo ratings yet
- Penang Matriculation College SB015 2018/2019: By: (MS1813362299 / H3P02B)Document11 pagesPenang Matriculation College SB015 2018/2019: By: (MS1813362299 / H3P02B)Yi LingNo ratings yet
- DNA ReplicationDocument7 pagesDNA Replicationlubuto TuntepeNo ratings yet
- DNA Synthesis: Further ReadingDocument1 pageDNA Synthesis: Further ReadingLaila Kurnia PramonoNo ratings yet
- 2.7 NewDocument64 pages2.7 NewReeya WhabiNo ratings yet
- Molecular Biology Unit IV and VDocument60 pagesMolecular Biology Unit IV and VchitraNo ratings yet
- CertificateDocument15 pagesCertificateAbhishek AryaNo ratings yet
- The Central Dogma of A Cell: DR S.Balakrishnan, PHD.Document58 pagesThe Central Dogma of A Cell: DR S.Balakrishnan, PHD.joyousNo ratings yet
- 6.2. DNA Replication PDFDocument75 pages6.2. DNA Replication PDFShania TehNo ratings yet
- DNA Replication and Protein SynthesisDocument5 pagesDNA Replication and Protein SynthesisFrezelVillaBasiloniaNo ratings yet
- 2 - Nucleic Acid Replication, Transcription & Translation - ModuleDocument15 pages2 - Nucleic Acid Replication, Transcription & Translation - ModuleMichaella CabanaNo ratings yet
- DNA's Structure and Function - CH 10Document10 pagesDNA's Structure and Function - CH 10jokersgang61No ratings yet
- DNA ReplicationDocument6 pagesDNA ReplicationGeorge TarraNo ratings yet
- DNAreplication Lehninger BiochemistryDocument14 pagesDNAreplication Lehninger BiochemistrySiddhant MazumderNo ratings yet
- Central Dogma of GeneticsDocument4 pagesCentral Dogma of Geneticsmiguel gaquitNo ratings yet
- Dna ReplicationDocument37 pagesDna ReplicationSyamala Natarajan100% (1)
- Replikasi DNA Dan Sintesis Protein (Gabungan)Document62 pagesReplikasi DNA Dan Sintesis Protein (Gabungan)adinda tyasprabandariNo ratings yet
- Presentationprint TempDocument27 pagesPresentationprint TempGandepelli SwetchaNo ratings yet
- Compressed Notes Chapter 6: Expression of Biological Information Sb015Document10 pagesCompressed Notes Chapter 6: Expression of Biological Information Sb015SYAZWAN BIN MUSTAFA MoeNo ratings yet
- Individual Assignment 1 PPH 226Document3 pagesIndividual Assignment 1 PPH 226Benjamin Aluoch OnyangoNo ratings yet
- DNA ReplicationDocument18 pagesDNA ReplicationBalamurali PandiyanNo ratings yet
- Central Dogma of LifeDocument17 pagesCentral Dogma of LifeSathish KumarNo ratings yet
- 2.7 DNA Replication, Transcription, and TranslationDocument46 pages2.7 DNA Replication, Transcription, and Translationpapa3rikusNo ratings yet
- Replication PDFDocument12 pagesReplication PDFYi LingNo ratings yet
- Molecular Biology: BBM FK UntarDocument47 pagesMolecular Biology: BBM FK UntarfaustineNo ratings yet
- All About DNA: Sci 211 - GeneticsDocument13 pagesAll About DNA: Sci 211 - GeneticsPATRICK MARK FLORESNo ratings yet
- The Growing Point Paradox and Discontinuous DNA SynthesisDocument5 pagesThe Growing Point Paradox and Discontinuous DNA Synthesisintan yunandaNo ratings yet
- Nucleic Acids NOTESDocument8 pagesNucleic Acids NOTESelaineNo ratings yet
- Dna ReplicationDocument28 pagesDna ReplicationavielavenderNo ratings yet
- DNA Replication, The Basis ForDocument23 pagesDNA Replication, The Basis ForDiana M. LlonaNo ratings yet
- 356 - Cell-Biology Physiology) DNA Replication001Document6 pages356 - Cell-Biology Physiology) DNA Replication001Leen AlhussainNo ratings yet
- DNA Structure and ReplicationDocument12 pagesDNA Structure and ReplicationJhune Dominique GalangNo ratings yet
- Nucleic Acids SummaryDocument4 pagesNucleic Acids SummaryIvan MiglioriniNo ratings yet
- Molecular Biology Lecture 10BDocument51 pagesMolecular Biology Lecture 10BAnton IrvingNo ratings yet
- Bisc207 Reviewer 6Document1 pageBisc207 Reviewer 6Anabelle McKenlyNo ratings yet
- 4 DnaDocument6 pages4 Dnadr_47839666No ratings yet
- Nucleic Acids - Topic 7 HLDocument28 pagesNucleic Acids - Topic 7 HLGracieNo ratings yet
- DNA ReplicationDocument28 pagesDNA ReplicationAsim Bin Arshad 58-FBAS/BSBIO/S19100% (1)
- Sanger Sequencing and The Polymerase Chain ReactionDocument4 pagesSanger Sequencing and The Polymerase Chain ReactionNicolas JayNo ratings yet
- 2-BT1000-Lec12 DNA Replication - IDocument14 pages2-BT1000-Lec12 DNA Replication - IShashwat SahooNo ratings yet
- Molecular Biology Study MaterialDocument58 pagesMolecular Biology Study MaterialchitraNo ratings yet
- 2.7 DNA Replication, Transcription and TranslationDocument46 pages2.7 DNA Replication, Transcription and TranslationAcyl HoteitNo ratings yet
- 2.7 7.1DNAReplicationDocument33 pages2.7 7.1DNAReplicationKhin (Darin) Hnin PhyuNo ratings yet
- Mechanisms of DNA Synthesis: Biochemistry 201 Advanced Molecular Biology January 19, 2000 Doug BrutlagDocument12 pagesMechanisms of DNA Synthesis: Biochemistry 201 Advanced Molecular Biology January 19, 2000 Doug BrutlagJessa CabañaNo ratings yet
- Dna Replication in ProkaryotesDocument10 pagesDna Replication in ProkaryotesLekshmyNo ratings yet
- 3.dna ReplicationDocument7 pages3.dna ReplicationCharm BatiancilaNo ratings yet
- DNA Replication: From Wikipedia, The Free EncyclopediaDocument10 pagesDNA Replication: From Wikipedia, The Free EncyclopediaRahul GuptaNo ratings yet
- Chap 7 Part 1 NUCLEIC ACIDSDocument37 pagesChap 7 Part 1 NUCLEIC ACIDSThe HushNo ratings yet
- Organization of Bacterial GenomesDocument24 pagesOrganization of Bacterial GenomeshariNo ratings yet
- Theoretical EcologyDocument4 pagesTheoretical EcologyFaieryNo ratings yet
- Adaptations in Vertebrate EvolutionDocument2 pagesAdaptations in Vertebrate EvolutionFaieryNo ratings yet
- He Measured All Live Trees in Plots TwiceDocument2 pagesHe Measured All Live Trees in Plots TwiceFaieryNo ratings yet
- Chapter 2: Conditions: Extra-Reading: Lectures 2 and 5Document8 pagesChapter 2: Conditions: Extra-Reading: Lectures 2 and 5FaieryNo ratings yet
- GLOMEROMYCOTADocument9 pagesGLOMEROMYCOTAFaieryNo ratings yet
- EE.06 ExtrareadingDocument6 pagesEE.06 ExtrareadingFaieryNo ratings yet
- Theoretical EcologyDocument4 pagesTheoretical EcologyFaieryNo ratings yet
- Adaptations in Vertebrate EvolutionDocument3 pagesAdaptations in Vertebrate EvolutionFaieryNo ratings yet
- Expectedoutside. CBG.02Document4 pagesExpectedoutside. CBG.02FaieryNo ratings yet
- ExpectedOutside. CBG.07Document8 pagesExpectedOutside. CBG.07FaieryNo ratings yet
- Glo Me Romy CotaDocument10 pagesGlo Me Romy CotaFaieryNo ratings yet
- Expectedoutside. CBG.02Document5 pagesExpectedoutside. CBG.02FaieryNo ratings yet
- ExpectedOutside. CBG.01Document10 pagesExpectedOutside. CBG.01FaieryNo ratings yet
- CBG 04Document2 pagesCBG 04FaieryNo ratings yet
- Twin StudiesDocument1 pageTwin StudiesFaieryNo ratings yet
- CBG 07Document1 pageCBG 07FaieryNo ratings yet
- Gene SeqDocument4 pagesGene SeqFaieryNo ratings yet
- Metabolism UlDocument1 pageMetabolism UlFaieryNo ratings yet
- H3 SmokingDocument1 pageH3 SmokingFaieryNo ratings yet
- Common BiochemicalsDocument7 pagesCommon BiochemicalsFaieryNo ratings yet
- Working GroupsDocument1 pageWorking GroupsFaieryNo ratings yet
- Is The World Changing For The BetterDocument2 pagesIs The World Changing For The BetterFaieryNo ratings yet
- Vocab UnitsDocument62 pagesVocab UnitsFaieryNo ratings yet
- Integrated WritingDocument5 pagesIntegrated WritingFaieryNo ratings yet
- DowsingDocument1 pageDowsingFaieryNo ratings yet
- ClovisDocument1 pageClovisFaieryNo ratings yet
- H1 TV ViolenceDocument1 pageH1 TV ViolenceFaieryNo ratings yet
- H2 - The Best TeacherDocument1 pageH2 - The Best TeacherFaieryNo ratings yet
- AsthmaDocument1 pageAsthmaFaieryNo ratings yet
- Autophagy PosterDocument1 pageAutophagy Postersonam_c100% (1)
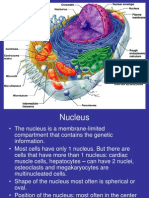
- NucleusDocument58 pagesNucleusImperiall100% (1)
- IB HL BIO FULL NOTES (Onarıldı)Document874 pagesIB HL BIO FULL NOTES (Onarıldı)Sıla DenizNo ratings yet
- Lecture 9-GS1 1112-Mitochondria & ChloroplastDocument37 pagesLecture 9-GS1 1112-Mitochondria & ChloroplastHa LuongNo ratings yet
- 03 Protein MetabolismDocument37 pages03 Protein MetabolismCedrix CuadernoNo ratings yet
- 9700 m18 QP 22 PDFDocument16 pages9700 m18 QP 22 PDFIG UnionNo ratings yet
- DR Simran Tandon - CV PDFDocument1 pageDR Simran Tandon - CV PDFSumeet GillNo ratings yet
- 13 - GluconeogenesisDocument23 pages13 - GluconeogenesischeckmateNo ratings yet
- BP504 T. (Pharmacognosy AND Phytochemistry II (Theory) )Document11 pagesBP504 T. (Pharmacognosy AND Phytochemistry II (Theory) )gaurav saxenaNo ratings yet
- Chapter 3 - Cell Structure & Functions-1 PDFDocument17 pagesChapter 3 - Cell Structure & Functions-1 PDFSTEM 8 PRODUCTIONSNo ratings yet
- Histology I: Dr. Mustafa Ghanim & Dr. Fatina HanbaliDocument35 pagesHistology I: Dr. Mustafa Ghanim & Dr. Fatina Hanbalijude dwekatNo ratings yet
- Jurnal Biosel Serial Endosymbiosis TheoryDocument4 pagesJurnal Biosel Serial Endosymbiosis TheoryHeny SinagaNo ratings yet
- Animal Tissue: Chapter - 10 Std. XIDocument110 pagesAnimal Tissue: Chapter - 10 Std. XIYash Soneji100% (2)
- Neurobiophysics2009 PreprintDocument52 pagesNeurobiophysics2009 PreprintsmiglemNo ratings yet
- Lecture at University of IndonesiaDocument58 pagesLecture at University of IndonesiaFikriNo ratings yet
- Bombay Blood GroupDocument19 pagesBombay Blood GroupchinnnababuNo ratings yet
- Soalan Objektif Mac2014Document2 pagesSoalan Objektif Mac2014Siti NajwaNo ratings yet
- 1 Resveratrol Internat Immunopharm2019Document13 pages1 Resveratrol Internat Immunopharm2019Mtro. Javier Alfredo Carballo PereaNo ratings yet
- List of Clotting FactorsDocument1 pageList of Clotting FactorsGNiqMNo ratings yet
- Microbial GrowthDocument48 pagesMicrobial Growthdreamrose100% (14)
- In Situ Bio Printing of The Skin For BurnsDocument1 pageIn Situ Bio Printing of The Skin For BurnsJean-Philippe RivièreNo ratings yet
- Stahl and Meselson'S Work On Semiconservative Nature of Dna ReplicationDocument12 pagesStahl and Meselson'S Work On Semiconservative Nature of Dna ReplicationKima MadNo ratings yet
- Cytogenetics: Preliminaries: Mccarty Shows That Dna Can Transform BacteriaDocument5 pagesCytogenetics: Preliminaries: Mccarty Shows That Dna Can Transform BacteriaRomie SolacitoNo ratings yet
- C Science 10 Quarter 3 Module 3 (Week 4)Document14 pagesC Science 10 Quarter 3 Module 3 (Week 4)Daisy Soriano Prestoza100% (1)
- MainDocument13 pagesMainIkmah FauzanNo ratings yet