You might also like
- DD-transpeptidase: From, The Free EncyclopediaDocument1 pageDD-transpeptidase: From, The Free EncyclopediaAndreas StavrianosNo ratings yet
- Analysis of Rab GTPases in Dictyostelium discoideumDocument34 pagesAnalysis of Rab GTPases in Dictyostelium discoideumdavid_stephens_29No ratings yet
- ChaperoneDocument13 pagesChaperoneRomana MasnikosaNo ratings yet
- Mol Bio Part 1 2014Document95 pagesMol Bio Part 1 2014Manish SoniNo ratings yet
- Zamow Art PDF - PHTMLDocument17 pagesZamow Art PDF - PHTMLمحمد نعيمNo ratings yet
- Improvement and Also Consent of An Basic Oriental Model Plus A FaceScale Type of Your Oxford Neck Credit Score The 2center Prospective Examinexsbvy PDFDocument1 pageImprovement and Also Consent of An Basic Oriental Model Plus A FaceScale Type of Your Oxford Neck Credit Score The 2center Prospective Examinexsbvy PDFmoatcause47No ratings yet
- A Conserved Arginine Plays A Role in The Catalytic Cycle of The Protein Disulphide IsomerasesDocument13 pagesA Conserved Arginine Plays A Role in The Catalytic Cycle of The Protein Disulphide Isomeraseshiran.reisNo ratings yet
- Science Glycerol 5Document11 pagesScience Glycerol 5Iju IzzuNo ratings yet
- Cell-free Protein Synthesis of Membrane (1,3) -Β-D-glucan (Curdlan) SynthaseDocument15 pagesCell-free Protein Synthesis of Membrane (1,3) -Β-D-glucan (Curdlan) SynthasecesamavNo ratings yet
- Nucleic AcidDocument88 pagesNucleic AcidJoyce Jimenez100% (1)
- Low-Resolution Structure of Drosophila Translin: Vinay Kumar, Gagan D. GuptaDocument10 pagesLow-Resolution Structure of Drosophila Translin: Vinay Kumar, Gagan D. GuptaGagan D GuptaNo ratings yet
- Enzyme Mech ExamplesDocument9 pagesEnzyme Mech ExamplesJosé Rafael TovarNo ratings yet
- The Mechanism of Action of Penicillin: ProcDocument10 pagesThe Mechanism of Action of Penicillin: Procadriani lestariNo ratings yet
- Eukaryotic DNA Polymerases: Sue Cotterill, Stephen KearseyDocument6 pagesEukaryotic DNA Polymerases: Sue Cotterill, Stephen KearseySoma GhoshNo ratings yet
- Dna Genes Chromosomes 2011Document65 pagesDna Genes Chromosomes 2011Suliman GarallehNo ratings yet
- Carey 1997 PDFDocument16 pagesCarey 1997 PDFCsilla NémethNo ratings yet
- Crystal Structure of A Bacterial Signal Peptidase ApoenzymeDocument8 pagesCrystal Structure of A Bacterial Signal Peptidase ApoenzymeOktavia firnandaNo ratings yet
- DGR Literature ReviewDocument2 pagesDGR Literature ReviewIvaNo ratings yet
- 02 Paper 05 Michael ThreadgillDocument14 pages02 Paper 05 Michael ThreadgillOmar Nassir MoftahNo ratings yet
- Paper - IV Molecular BiologyDocument236 pagesPaper - IV Molecular BiologyNaruto UzumakiNo ratings yet
- Naem:-Mohammed Naem يويلع ميعن دمحم -:مسلاا University:- ISUCOD 2 Stage Subject:- cathepsin D Teacher master:Document10 pagesNaem:-Mohammed Naem يويلع ميعن دمحم -:مسلاا University:- ISUCOD 2 Stage Subject:- cathepsin D Teacher master:محمد نعيمNo ratings yet
- Tutorial 3 - Biology 101 Answer MemoDocument18 pagesTutorial 3 - Biology 101 Answer MemoKaizer NdoloNo ratings yet
- D-Xylose 1-Dehydrogenase: Jump To Navigationjump To SearchDocument2 pagesD-Xylose 1-Dehydrogenase: Jump To Navigationjump To SearchLuis Angel Flores CordovaNo ratings yet
- Mode of Action of Vancomycin: L D D D D DDocument8 pagesMode of Action of Vancomycin: L D D D D DNolanNo ratings yet
- Karimi 2020Document13 pagesKarimi 2020Nelson Daniel Marcano AguileraNo ratings yet
- European Journal of Biochemistry - 2002 - Cherepanov - Dynamic Mechanism of Nick Recognition by DNA LigaseDocument7 pagesEuropean Journal of Biochemistry - 2002 - Cherepanov - Dynamic Mechanism of Nick Recognition by DNA LigaseAnuvansh SinghNo ratings yet
- Escherichia Coli DH10BDocument2 pagesEscherichia Coli DH10BSecc. 2 Marco Antonio Aviles RomeroNo ratings yet
- Nucleic Acid Structure and FunctionDocument30 pagesNucleic Acid Structure and FunctionZayan HaiderNo ratings yet
- 1989 Hanukoglu Gutfinger European Journal of Biochemistry FEBS 180 2 cDNA Sequence of Adrenodoxin ReductaseDocument6 pages1989 Hanukoglu Gutfinger European Journal of Biochemistry FEBS 180 2 cDNA Sequence of Adrenodoxin Reductasel4vfeaokf5No ratings yet
- Best Practice & Research Clinical Endocrinology & MetabolismDocument14 pagesBest Practice & Research Clinical Endocrinology & MetabolismF Aurelia AnndNo ratings yet
- BiochemDocument383 pagesBiochemtylermedNo ratings yet
- P450-Mediated Carbon–Carbon Ring Closure in Lathyrane Diterpenoid BiosynthesisDocument5 pagesP450-Mediated Carbon–Carbon Ring Closure in Lathyrane Diterpenoid BiosynthesisEdithNo ratings yet
- RNA Binding ProteinDocument8 pagesRNA Binding ProteinBhagyaNo ratings yet
- Helicase Enzyme: BiochemistryDocument9 pagesHelicase Enzyme: BiochemistryTayf AlrawINo ratings yet
- Mila NitipDocument6 pagesMila NitipMulyati SafiraNo ratings yet
- Word ListDocument14 pagesWord ListShubhamNo ratings yet
- Paper 4 QuickiesDocument40 pagesPaper 4 QuickiesRobin KeaneNo ratings yet
- Nucleic AcidsDocument20 pagesNucleic AcidsIsrat Jahan SurovyNo ratings yet
- Escherichia coli DH10B (2)Document2 pagesEscherichia coli DH10B (2)Carlos Alberto Alva SifuentesNo ratings yet
- Cosenza 2002 PDFDocument17 pagesCosenza 2002 PDFLarry CosenzaNo ratings yet
- Primeau Et Al-2011-FEBS LettersDocument7 pagesPrimeau Et Al-2011-FEBS LettersSergeat18BNo ratings yet
- Xie 2015. Diacylglycerol Kinases in Membrane TraffickingDocument10 pagesXie 2015. Diacylglycerol Kinases in Membrane TraffickingmcrickboomNo ratings yet
- VBDFGHVCVSDGDocument8 pagesVBDFGHVCVSDGajayNo ratings yet
- Proteolytic Enzyme: Basic Information and Cleavage Rules: Dr. Aditya AryaDocument11 pagesProteolytic Enzyme: Basic Information and Cleavage Rules: Dr. Aditya Aryaabcxyz7799No ratings yet
- Conversion of Nucleotides Sequences Into Genomic SignalsDocument25 pagesConversion of Nucleotides Sequences Into Genomic SignalsChandrasekaran SaravanakumarNo ratings yet
- Nucleic Acids Tutorial - DNA, RNA Structure & Function (40Document20 pagesNucleic Acids Tutorial - DNA, RNA Structure & Function (40Yen Wei goNo ratings yet
- Lecture 26: Enzyme Mechanism: Serine Proteases: Biochemistry I Fall Term, 2004Document5 pagesLecture 26: Enzyme Mechanism: Serine Proteases: Biochemistry I Fall Term, 2004Louis FortunatoNo ratings yet
- Kotov 2014 EnglDocument13 pagesKotov 2014 EnglLudmila OleninaNo ratings yet
- Sonnenburg Biochemistry2003Document11 pagesSonnenburg Biochemistry2003Claudia BuitragoNo ratings yet
- Siezen, 1997, SubtilaseDocument23 pagesSiezen, 1997, SubtilaseElodie LefebvreNo ratings yet
- Nucleic acid study notes - Fall 2018Document10 pagesNucleic acid study notes - Fall 2018ReggieNo ratings yet
- Aji Sutrisno Food Science and Technology Universitas Brawijaya (Ub)Document81 pagesAji Sutrisno Food Science and Technology Universitas Brawijaya (Ub)Nirma MardinitaNo ratings yet
- Aji Sutrisno Food Science and Technology Universitas Brawijaya (Ub)Document81 pagesAji Sutrisno Food Science and Technology Universitas Brawijaya (Ub)Gandhes Cintya DewiNo ratings yet
- Origin Remodeling and Opening in Bacteria Rely On Distinct Assembly States of The Dnaa InitiatorDocument11 pagesOrigin Remodeling and Opening in Bacteria Rely On Distinct Assembly States of The Dnaa InitiatorDina A. ŠabićNo ratings yet
- Alkaline Lysis, SDS Page, Sequencing, ChromatographyDocument73 pagesAlkaline Lysis, SDS Page, Sequencing, ChromatographyShashwat Mishra100% (1)
- QI II - Platinum Coordination Compounds With Potent Anticancer ActivityDocument17 pagesQI II - Platinum Coordination Compounds With Potent Anticancer ActivityAbel de Jesús RiveraNo ratings yet
- DNA Structure RevealedDocument44 pagesDNA Structure Revealedevacarlina1721No ratings yet
- Science:, 1040 (2013) Helen R. SaibilDocument3 pagesScience:, 1040 (2013) Helen R. SaibilPablo Antonio Galaz DavisonNo ratings yet
- DNA Rep and RNA StructureDocument30 pagesDNA Rep and RNA Structurekoromamoses235No ratings yet
- Alanine RacemaseDocument6 pagesAlanine RacemaseHerdiyan Yogi SugaraNo ratings yet
- Prosedur RheologiDocument1 pageProsedur RheologiHerdiyan Yogi SugaraNo ratings yet
- DD-transpeptidase: Bacterial enzyme targeted by penicillinDocument4 pagesDD-transpeptidase: Bacterial enzyme targeted by penicillinHerdiyan Yogi SugaraNo ratings yet
- Inositol MonophosphataseDocument5 pagesInositol MonophosphataseHerdiyan Yogi SugaraNo ratings yet
- Isi No 3 Hasib-IrmaDocument4 pagesIsi No 3 Hasib-IrmaHerdiyan Yogi SugaraNo ratings yet
- R. Landolt 1. (-) Polivalen (Tidak Ada Endapan) (+) Monovalen (Endapan Putih)Document1 pageR. Landolt 1. (-) Polivalen (Tidak Ada Endapan) (+) Monovalen (Endapan Putih)Herdiyan Yogi SugaraNo ratings yet
- Let Her Go - Passenger: Need The LightDocument2 pagesLet Her Go - Passenger: Need The LightakuiniaNo ratings yet
- Borobudur TempleDocument1 pageBorobudur TempleHerdiyan Yogi SugaraNo ratings yet
- ACE Biology O'level Book@2021Document213 pagesACE Biology O'level Book@2021DuyNo ratings yet
- The Importance of Anaerobic ExerciseDocument13 pagesThe Importance of Anaerobic Exerciseapi-357421918No ratings yet
- Selection of Artificial TeethDocument44 pagesSelection of Artificial TeethMohamed VandettaNo ratings yet
- Respiratory Muscle Ultrasonography: Methodology, Basic and Advanced Principles and Clinical Applications in ICU and ED Patients-A Narrative ReviewDocument12 pagesRespiratory Muscle Ultrasonography: Methodology, Basic and Advanced Principles and Clinical Applications in ICU and ED Patients-A Narrative Reviewalejandro RodriguezNo ratings yet
- 0010 (1) Science - Terms 1.154KB PDFDocument236 pages0010 (1) Science - Terms 1.154KB PDFMuhammad SaleemNo ratings yet
- Nutrition Folio (Basal Metabolic Rate)Document15 pagesNutrition Folio (Basal Metabolic Rate)Nursakinah NajwahNo ratings yet
- Pro To Zoology 1954 KudoDocument988 pagesPro To Zoology 1954 KudoClaau GomezNo ratings yet
- Trimestral Exam First GradeDocument3 pagesTrimestral Exam First Gradeemmanuel espinozaNo ratings yet
- Essentials Lab Resource Guide 12 22Document67 pagesEssentials Lab Resource Guide 12 22Aijeleth Shahar Gunay AwacayNo ratings yet
- Use of Vasopressors and Inotropes - UpToDateDocument25 pagesUse of Vasopressors and Inotropes - UpToDateVictor Mendoza - MendezNo ratings yet
- MSDS Garam MejaDocument5 pagesMSDS Garam MejaDesyrulaNo ratings yet
- PTT 311 (Assignment 2) (Answer Script)Document4 pagesPTT 311 (Assignment 2) (Answer Script)Berkcan ArslanNo ratings yet
- Interval Training For PerformanceDocument20 pagesInterval Training For PerformancePaulo Tsuneta100% (1)
- Human Motivation Edited2Document23 pagesHuman Motivation Edited2jingoNo ratings yet
- Medication AdministrationDocument88 pagesMedication AdministrationKBD100% (1)
- Microtome and ultrastructure microscopy guideDocument360 pagesMicrotome and ultrastructure microscopy guidesquishyboitae33% (3)
- Essencial Phrasal VerbsDocument4 pagesEssencial Phrasal VerbsalvereNo ratings yet
- Part of Brain 2Document2 pagesPart of Brain 2PonCut Teuku AchyarNo ratings yet
- Regenesis 1Document14 pagesRegenesis 1White Light100% (3)
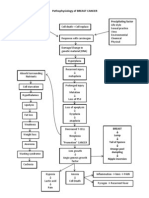
- Pathophysiology of BREAST CANCERDocument1 pagePathophysiology of BREAST CANCERAlinor Abubacar100% (6)
- Ineffective Airway ClearanceDocument1 pageIneffective Airway ClearanceFreisanChenMandumotanNo ratings yet
- Kantor Cabang: BANJARMASIN - 1701 FKTP: Kertak Hanyar - 17040601Document6 pagesKantor Cabang: BANJARMASIN - 1701 FKTP: Kertak Hanyar - 17040601Ic-tika Siee ChuabbieNo ratings yet
- Q and A DactylosDocument56 pagesQ and A DactylosJUNN REE MONTILLA100% (2)
- Test Bank For Mechanical Ventilation 7th Edition J M CairoDocument11 pagesTest Bank For Mechanical Ventilation 7th Edition J M CairoJohnCampbellyacer100% (27)
- Indications For Initiation of DialysisDocument11 pagesIndications For Initiation of DialysisMilton BenevidesNo ratings yet
- Physiology of The Liver: Corresponding AuthorDocument12 pagesPhysiology of The Liver: Corresponding AuthorMansour HazaNo ratings yet
- Chronic Cough Differential DiagnosisDocument6 pagesChronic Cough Differential DiagnosisUbaidillah HafidzNo ratings yet
- Preparing Blood ComponentsDocument9 pagesPreparing Blood ComponentsaksinuNo ratings yet
- Asthma Care Quick Reference - Diagnosing and Managing AsthmaDocument18 pagesAsthma Care Quick Reference - Diagnosing and Managing AsthmaMarizka Putri AftriaNo ratings yet
- Arjun Chauhan (Bio)Document52 pagesArjun Chauhan (Bio)Sharad ChoudharyNo ratings yet