You might also like
- Research Article: Composition of Algal Oil and Its Potential As BiofuelDocument15 pagesResearch Article: Composition of Algal Oil and Its Potential As BiofuelTeresa MataNo ratings yet
- Microalgal Co-Cultivation For Biofuel Production and Bioremediation: Current Status and BenefitsDocument26 pagesMicroalgal Co-Cultivation For Biofuel Production and Bioremediation: Current Status and BenefitsJonathas FerreiraNo ratings yet
- Advancement of Green Technologies - A Comprehensive Review On The Potential Application of Microalgae BiomassDocument15 pagesAdvancement of Green Technologies - A Comprehensive Review On The Potential Application of Microalgae Biomassbambang_teknikkimiaNo ratings yet
- Novel Stacked Modular Open Raceway Ponds For Microalgae Biomass Cultivation in Biogas Plants: Preliminary Design and ModellingDocument19 pagesNovel Stacked Modular Open Raceway Ponds For Microalgae Biomass Cultivation in Biogas Plants: Preliminary Design and ModellingFábio UcellaNo ratings yet
- Pre-Treatment Methods For Production of Biofuel From Microalgae BiomassDocument12 pagesPre-Treatment Methods For Production of Biofuel From Microalgae Biomassrian arisandiNo ratings yet
- Biomass From Microalgae: The Potential of Domestication Towards Sustainable BiofactoriesDocument18 pagesBiomass From Microalgae: The Potential of Domestication Towards Sustainable BiofactoriesggarzonjNo ratings yet
- Progress of Microalgae Biofuels CommerciDocument10 pagesProgress of Microalgae Biofuels CommerciNthambi MiriamNo ratings yet
- Biofuels From Microalgae PDFDocument6 pagesBiofuels From Microalgae PDFluis APNo ratings yet
- Tredici Photobiology 2010Document20 pagesTredici Photobiology 2010Luis EspinozaNo ratings yet
- Algae Biomass As A Potential Source of Liquid FuelsDocument14 pagesAlgae Biomass As A Potential Source of Liquid FuelsFranzhean Balais CuachonNo ratings yet
- AlgasDocument25 pagesAlgasDayanneNo ratings yet
- Biodiesel From Microalgae Environmental AspectsDocument4 pagesBiodiesel From Microalgae Environmental AspectsNaty RubilarNo ratings yet
- Walterborgesdeoliveira2011 PDFDocument11 pagesWalterborgesdeoliveira2011 PDFJairo ArenasNo ratings yet
- Enamala2018 PDFDocument20 pagesEnamala2018 PDFBriand VallejoNo ratings yet
- 8 Jmea E20130213 3Document11 pages8 Jmea E20130213 3Tichman MetrixNo ratings yet
- Microalgae Biofuel Review: Present & Future OutlookDocument10 pagesMicroalgae Biofuel Review: Present & Future OutlookAyesha MukhtarNo ratings yet
- Impact of Abiotic Factors On Biodiesel Production by MicroalgaeDocument11 pagesImpact of Abiotic Factors On Biodiesel Production by MicroalgaeSayuri GuevaraNo ratings yet
- Impact of Abiotic Factors On Biodiesel Production by MicroalgaeDocument11 pagesImpact of Abiotic Factors On Biodiesel Production by MicroalgaeSayuri GuevaraNo ratings yet
- Collet 2014Document9 pagesCollet 2014Monica AntolinezNo ratings yet
- Mass Cultivation and Harvesting of Microalgal Biomass: Current Trends and Future PerspectivesDocument14 pagesMass Cultivation and Harvesting of Microalgal Biomass: Current Trends and Future PerspectivesggarzonjNo ratings yet
- Journal of Cleaner Production Comparative Study 1-S2.0-S0959652622048107-MainDocument14 pagesJournal of Cleaner Production Comparative Study 1-S2.0-S0959652622048107-MainOmar KHELILNo ratings yet
- Para Quo SummaDocument7 pagesPara Quo SummaJarred Allen Z. ServañaNo ratings yet
- Food Industries Wastewater Recycling For Biodiesel Production Through Microalgal RemediationDocument17 pagesFood Industries Wastewater Recycling For Biodiesel Production Through Microalgal RemediationUmme HanieNo ratings yet
- Pretratamiento de Residuos Lignocelulosicos para La Obtencion de BiocombustiblesDocument15 pagesPretratamiento de Residuos Lignocelulosicos para La Obtencion de BiocombustiblesElena FloresNo ratings yet
- Algal biofuel challenges and prospectsDocument10 pagesAlgal biofuel challenges and prospectsRahul KumarNo ratings yet
- Renewable and Sustainable Energy Reviews: Laurencas Raslavi Čius, Nerijus Stri Ūgas, Mantas FelnerisDocument12 pagesRenewable and Sustainable Energy Reviews: Laurencas Raslavi Čius, Nerijus Stri Ūgas, Mantas FelnerisGirard Immanuel SorianoNo ratings yet
- Fuels. Fuels Fuels: Stern, 2006Document7 pagesFuels. Fuels Fuels: Stern, 2006annjie99No ratings yet
- Curs Biogaz - 4075Document22 pagesCurs Biogaz - 4075Ana-Maria PopescuNo ratings yet
- 2011 - Stephenson Et Al. - Improving Photosynthesis For Algal Biofuels Toward A Green RevolutionDocument9 pages2011 - Stephenson Et Al. - Improving Photosynthesis For Algal Biofuels Toward A Green RevolutionmbrancovNo ratings yet
- Biotechnology Reports: ReviewDocument7 pagesBiotechnology Reports: ReviewdatinovNo ratings yet
- Haba Shy 2021Document15 pagesHaba Shy 2021nataliaimeldawNo ratings yet
- Energia Aplicada A BiorreactoresDocument40 pagesEnergia Aplicada A BiorreactoresMarianNo ratings yet
- E3sconf Eems2017 02032Document10 pagesE3sconf Eems2017 02032ali abdulrahman al-ezziNo ratings yet
- Ees 2016 0132Document17 pagesEes 2016 0132GREENLAB ECL TECHNo ratings yet
- A Review On Algae Biodiesel A Novel Source of Renewable Energy in IndiaDocument3 pagesA Review On Algae Biodiesel A Novel Source of Renewable Energy in IndiaEditor IJRITCCNo ratings yet
- Water Use and Its Recycling in Microalgae Cultivation For Biofuel ApplicationDocument9 pagesWater Use and Its Recycling in Microalgae Cultivation For Biofuel ApplicationStefano CarreñoNo ratings yet
- Cultivation Systems of Microalgae For The Production of BiofuelsDocument20 pagesCultivation Systems of Microalgae For The Production of BiofuelsRobson SantiagoNo ratings yet
- Rajabihamedani 2019Document11 pagesRajabihamedani 2019M Yousaf YaqoobNo ratings yet
- Bioresource Technology: D. Hernández, B. Riaño, M. Coca, M.C. García-GonzálezDocument6 pagesBioresource Technology: D. Hernández, B. Riaño, M. Coca, M.C. García-GonzálezAndres MilquezNo ratings yet
- Parmar2011 PDFDocument10 pagesParmar2011 PDFsaima afridiNo ratings yet
- Alok Kumar Singh, Et AlDocument11 pagesAlok Kumar Singh, Et AlKavinNo ratings yet
- Algal Research: Review ArticleDocument19 pagesAlgal Research: Review ArticleXuan NguyenNo ratings yet
- Applications of Microalgal Biofilms For WastewaterDocument24 pagesApplications of Microalgal Biofilms For WastewaterTrần T. MơNo ratings yet
- Cherubini - Crop Residues As Raw Materials For Biorefinery Systems - A LCA Case StudyDocument11 pagesCherubini - Crop Residues As Raw Materials For Biorefinery Systems - A LCA Case StudyGuiping HuNo ratings yet
- Microalgae To Biofuels Promising' Alternative and Renewable EnergyDocument13 pagesMicroalgae To Biofuels Promising' Alternative and Renewable EnergyJeevan KumarNo ratings yet
- Biorreactor: Un Sistema de Soporte BiológicoDocument8 pagesBiorreactor: Un Sistema de Soporte BiológicoDullahanovNo ratings yet
- A Comprehensive Review On Thermochemical, and Biochemical Conversion Methods of Lignocellulosic Biomass Into Valuable End ProductDocument9 pagesA Comprehensive Review On Thermochemical, and Biochemical Conversion Methods of Lignocellulosic Biomass Into Valuable End ProductAndreNo ratings yet
- GCB Bioenergy - 2014 - Yuan - Mass Balance and Life Cycle Assessment of Biodiesel From Microalgae Incorporated WithDocument15 pagesGCB Bioenergy - 2014 - Yuan - Mass Balance and Life Cycle Assessment of Biodiesel From Microalgae Incorporated WithitbwngNo ratings yet
- Bioresource Technology: Wasif Farooq, William I. Suh, Min S. Park, Ji-Won YangDocument9 pagesBioresource Technology: Wasif Farooq, William I. Suh, Min S. Park, Ji-Won YangKeli SobralNo ratings yet
- 2020 - Rajesh Banu Et Al. - Microalgae Based Biorefinery Promoting Circular Bioeconomy-Techno Economic and Life-Cycle AnalysisDocument14 pages2020 - Rajesh Banu Et Al. - Microalgae Based Biorefinery Promoting Circular Bioeconomy-Techno Economic and Life-Cycle AnalysismbrancovNo ratings yet
- Energy Conversion and Management: XDocument10 pagesEnergy Conversion and Management: XSagar RathodNo ratings yet
- (I) Chew 2017. Microalgae BiorefineryDocument10 pages(I) Chew 2017. Microalgae BiorefinerySacra PsyntergiaNo ratings yet
- Jurnal Bioetanol MikroalgaDocument22 pagesJurnal Bioetanol MikroalgaAkuf Suradal WibisonoNo ratings yet
- NG Et Al 2017Document13 pagesNG Et Al 2017rinifiahNo ratings yet
- 05 Aextj 366 23Document24 pages05 Aextj 366 23BRNSS Publication Hub InfoNo ratings yet
- Microalgal Drying and Cell Disruption - Recent AdvancesDocument9 pagesMicroalgal Drying and Cell Disruption - Recent AdvancesJuan Diego CárdenasNo ratings yet
- Applied SciencesDocument24 pagesApplied SciencesIonut NistorNo ratings yet
- Dębowski Et Al., 2022 ALGASDocument15 pagesDębowski Et Al., 2022 ALGASmaczweiNo ratings yet
- Utilization of Microalgal Bacterial Energy Nexus Improves - 2023 - EcotoxicologyDocument16 pagesUtilization of Microalgal Bacterial Energy Nexus Improves - 2023 - EcotoxicologyCristhian RodriguezNo ratings yet
- Commissioning Report 24V: Instruction PageDocument8 pagesCommissioning Report 24V: Instruction PageAngel Quijano ArmengolNo ratings yet
- Ultra-low maintenance Vantage batteries deliver 20+ years of reliable backup powerDocument8 pagesUltra-low maintenance Vantage batteries deliver 20+ years of reliable backup powerAngel Quijano ArmengolNo ratings yet
- ALCAD L - M - H Ranges: Nickel Cadmium BatteriesDocument2 pagesALCAD L - M - H Ranges: Nickel Cadmium BatteriesAngel Quijano ArmengolNo ratings yet
- Instalacion y OperacionDocument2 pagesInstalacion y OperacionAngel Quijano ArmengolNo ratings yet
- Vantage: Ultra-Low Maintenance BatteriesDocument8 pagesVantage: Ultra-Low Maintenance BatteriesAngel Quijano ArmengolNo ratings yet
- Light ScatteringDocument3 pagesLight ScatteringAngel Quijano ArmengolNo ratings yet
- Single Cell RangeDocument27 pagesSingle Cell RangeAngel Quijano ArmengolNo ratings yet
- Vantage: Installation and Operating InstructionsDocument2 pagesVantage: Installation and Operating InstructionsAngel Quijano ArmengolNo ratings yet
- Instalacion y OperacionDocument2 pagesInstalacion y OperacionAngel Quijano ArmengolNo ratings yet
- Biodegradation of Diesel Oil in Soil - 1Document8 pagesBiodegradation of Diesel Oil in Soil - 1Angel Quijano ArmengolNo ratings yet
- Pro II Input ManualDocument69 pagesPro II Input ManualNareshNo ratings yet
- Pro II 8.0 User GuideDocument390 pagesPro II 8.0 User GuideAngel Quijano ArmengolNo ratings yet
- Chapter-1 Nature of Crude PetroleumDocument7 pagesChapter-1 Nature of Crude PetroleumMahesh Kumar GuddeliNo ratings yet
- Nanotechnology For Algal BiofuelsDocument17 pagesNanotechnology For Algal BiofuelsAngel Quijano ArmengolNo ratings yet
- Absorption Spectra and Photovoltaic Characterization of Chlorophyllins PDFDocument8 pagesAbsorption Spectra and Photovoltaic Characterization of Chlorophyllins PDFAngel Quijano ArmengolNo ratings yet
- Ef 5000326Document6 pagesEf 5000326Angel Quijano ArmengolNo ratings yet
- Physiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamDocument61 pagesPhysiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamZaigham HammadNo ratings yet
- Adolescent Reproductive and Sexual HealthDocument42 pagesAdolescent Reproductive and Sexual HealthMuhammad Abbas WaliNo ratings yet
- Biology Investigatory ProjectDocument17 pagesBiology Investigatory ProjectAnmol Dhungel100% (6)
- MYCOVIRODocument11 pagesMYCOVIROPau SorianoNo ratings yet
- Behaviour of Great Apes Studied by Three Female ScientistsDocument12 pagesBehaviour of Great Apes Studied by Three Female ScientistsSani AhmadNo ratings yet
- 4546-LacOperon InstsDocument14 pages4546-LacOperon InstsScribd_is_GreatNo ratings yet
- 2010 FulltextDocument410 pages2010 FulltextNajam Din SahitoNo ratings yet
- Ssac 2104Document100 pagesSsac 2104Amrutha amrNo ratings yet
- MR Spectroscopy: Technical Aspects and Clinical ApplicationsDocument5 pagesMR Spectroscopy: Technical Aspects and Clinical ApplicationsNotariana Kusuma ANo ratings yet
- Trunking CatalogDocument8 pagesTrunking CatalogDenuka PathiranaNo ratings yet
- The "Five Families" College Essay ExampleDocument1 pageThe "Five Families" College Essay ExampleKishor RaiNo ratings yet
- 8 Cell - The Unit of Life-NotesDocument6 pages8 Cell - The Unit of Life-NotesBhavanya RavichandrenNo ratings yet
- Young Children's Biological Predisposition To Learn in Privileged DomainDocument6 pagesYoung Children's Biological Predisposition To Learn in Privileged DomainVeronica Dadal0% (1)
- Classification and Characterization of Microsatellite Instability Across 18 Cancer TypesDocument13 pagesClassification and Characterization of Microsatellite Instability Across 18 Cancer TypeshadymatrixNo ratings yet
- Mengenali Konflik Dalam Negosiasi Oleh: Zumaeroh: PendahuluanDocument47 pagesMengenali Konflik Dalam Negosiasi Oleh: Zumaeroh: PendahuluanrahmatNo ratings yet
- Lesson Plan No 5Document6 pagesLesson Plan No 5hafeez ahmedNo ratings yet
- Significance of 4D Printing For Dentistry: Materials, Process, and PotentialsDocument9 pagesSignificance of 4D Printing For Dentistry: Materials, Process, and PotentialsMohamed FaizalNo ratings yet
- Microbiological Test of Medical DevicesDocument5 pagesMicrobiological Test of Medical Devicesbijendra_sinhaNo ratings yet
- Week 6. Definision and DictionaryDocument19 pagesWeek 6. Definision and DictionaryafdreliautariazizahNo ratings yet
- IN SEARCH OF SEXUAL ENHANCEMENT THROUGH PILLS AND DRUGSDocument16 pagesIN SEARCH OF SEXUAL ENHANCEMENT THROUGH PILLS AND DRUGSJakob AndradeNo ratings yet
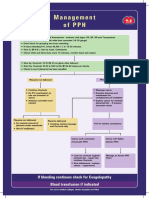
- Management of PPHDocument1 pageManagement of PPH098 U.KARTHIK SARAVANA KANTHNo ratings yet
- Plant Transport - IGCSE Biology Notes (2020)Document6 pagesPlant Transport - IGCSE Biology Notes (2020)Zhi En LeeNo ratings yet
- Chemical Composition of Meat and Nut Water of Major Coconut in PakistanDocument7 pagesChemical Composition of Meat and Nut Water of Major Coconut in PakistanSkyle VoNo ratings yet
- Regular Evaluation Test I Maths+Sc IXDocument7 pagesRegular Evaluation Test I Maths+Sc IXB.L. Indo Anglian Public School, Aurngabad, BiharNo ratings yet
- Muscle Energy Techniques ExplainedDocument2 pagesMuscle Energy Techniques ExplainedAtanu Datta100% (1)
- Vanders Human Physiology The Mechanisms of Body Function 15th Edition Widmaier Test BankDocument29 pagesVanders Human Physiology The Mechanisms of Body Function 15th Edition Widmaier Test BankDavidWardrcobi100% (15)
- Physiological Psychology ReviewerDocument13 pagesPhysiological Psychology ReviewerLeopando Rod CyrenzNo ratings yet
- Summary of Qualitative Tests (Pharmacognosy)Document8 pagesSummary of Qualitative Tests (Pharmacognosy)kidsaintfineNo ratings yet
- Student Exploration: Building DNADocument4 pagesStudent Exploration: Building DNAMia Smith100% (1)
- Cell Membrane and Transport ColoringDocument3 pagesCell Membrane and Transport ColoringTeresa GonzNo ratings yet