You might also like
- Esquema CatabolismoDocument10 pagesEsquema CatabolismoBegoña FolNo ratings yet
- Inhibidor EnzimáticoDocument18 pagesInhibidor EnzimáticoAdri CruzNo ratings yet
- Fosforilación OxidativaDocument4 pagesFosforilación OxidativaDaleska C. GalarzaNo ratings yet
- Enfermedades de Deposito LisosomalDocument24 pagesEnfermedades de Deposito LisosomalDaniel Coral100% (1)
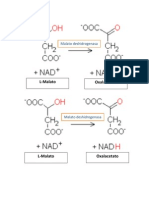
- La Enzima Malato DeshidrogenasaDocument2 pagesLa Enzima Malato DeshidrogenasaKaren Torres JuárezNo ratings yet
- Resumen de La Señalización CelularDocument22 pagesResumen de La Señalización CelularAndrésNo ratings yet
- Biosíntesis de Acidos GrasosDocument2 pagesBiosíntesis de Acidos Grasosbryer92No ratings yet
- Taller 2 - Transc Traducción.2019Document20 pagesTaller 2 - Transc Traducción.2019LOL X Repleys ElmoshtaykersNo ratings yet
- Jugos GastricosDocument8 pagesJugos GastricosBryan Giovanny ArmentaNo ratings yet
- Componentes de La Cadena RespiratoriaDocument5 pagesComponentes de La Cadena RespiratoriaHéctor De La Cruz BenitesNo ratings yet
- Proceso de Replicación +transcripción+traducciónDocument3 pagesProceso de Replicación +transcripción+traducciónjm rcNo ratings yet
- Fosfatos de Alta EnergiaDocument7 pagesFosfatos de Alta Energiachek0spNo ratings yet
- Glucolisis y Ciclo de KrebsDocument25 pagesGlucolisis y Ciclo de KrebsgabrielNo ratings yet
- Procesamiento SensitivoDocument56 pagesProcesamiento SensitivoMario Catalán Faúndez100% (1)
- Guia de La Via de La PentosaDocument3 pagesGuia de La Via de La PentosaOrfinaMarroquinNo ratings yet
- CICLO DE KREBS FLDocument32 pagesCICLO DE KREBS FLGabriela CarmonaNo ratings yet
- Lipogenesis FinDocument16 pagesLipogenesis Fintlotzin09No ratings yet
- Los Venenos de La Cadena RespiratoriaDocument6 pagesLos Venenos de La Cadena RespiratoriaArmando Patiño100% (1)
- Monografia MicroDocument11 pagesMonografia MicroRuth Nuria Alexandra Holgado ChallcoNo ratings yet
- Retículo Endoplásmico Pi MZVDocument29 pagesRetículo Endoplásmico Pi MZVCool BoyLVNo ratings yet
- Enzimología ClínicaDocument36 pagesEnzimología ClínicaSergio Melendez100% (1)
- Sindrome de HunterDocument18 pagesSindrome de HunterFernando Callata CardenasNo ratings yet
- Resumen Manual Histología Del Tejido ÓseoDocument8 pagesResumen Manual Histología Del Tejido Óseoa.andalopezNo ratings yet
- Via Del Acido ShiquimicoDocument29 pagesVia Del Acido ShiquimicoAnonymous cOnoOsmSSPNo ratings yet
- TEMA 6 Cultivos CelularesDocument11 pagesTEMA 6 Cultivos Celularesmomac100% (1)
- Tema 2. - Vía de Las Pentosas FosfatoDocument9 pagesTema 2. - Vía de Las Pentosas FosfatoandrearosescarNo ratings yet
- Biosíntesis de Los AminoácidosDocument4 pagesBiosíntesis de Los Aminoácidosmagda borjas100% (1)
- Qué Es Un Enlace HemiacetalicoDocument3 pagesQué Es Un Enlace HemiacetalicoFlores Gamarra DeylisNo ratings yet
- RibozimasDocument3 pagesRibozimasSalvador Salcedo67% (3)
- FotosistemasDocument6 pagesFotosistemasGUEVARA GALVEZ LEIDYNo ratings yet
- Fase LuminosaDocument3 pagesFase LuminosaWensy Frankier Ochoa MonteroNo ratings yet
- Metabolismo Del GLUCÓGENODocument35 pagesMetabolismo Del GLUCÓGENOErick Javier Guzman SerranoNo ratings yet
- Ciclo Del Ácido TricarboxílicoDocument32 pagesCiclo Del Ácido TricarboxílicoDenissLopez100% (1)
- Endosomas y LisosomasDocument24 pagesEndosomas y LisosomasAurelia Gonzales Calle93% (14)
- Resumen MicrosDocument3 pagesResumen MicrosChristian Huerta100% (1)
- EicosanoidesDocument25 pagesEicosanoidesMark Vertiz CernaNo ratings yet
- Actividad Ciclo de KrebsDocument3 pagesActividad Ciclo de KrebsWalter RiosNo ratings yet
- PEROXISOMASDocument7 pagesPEROXISOMASSol GutierrezNo ratings yet
- Protistas 1Document77 pagesProtistas 1Rodrigo CarralNo ratings yet
- Infecciones OportunistasDocument64 pagesInfecciones Oportunistasjrvg2007@gmail.comNo ratings yet
- FOSFORILACION OXIDATIVA LenningerDocument40 pagesFOSFORILACION OXIDATIVA LenningerTiktoker Carl JohnsonNo ratings yet
- Via de Las PentosasDocument25 pagesVia de Las PentosasHKlaudyNo ratings yet
- Tema 10 Bioquimica IIDocument4 pagesTema 10 Bioquimica IIYolanda Chaves GuerreroNo ratings yet
- Dinotrofenol MitocondriasDocument6 pagesDinotrofenol MitocondriasOskrNo ratings yet
- Clase 8 Lipogenesis 2014 PDFDocument36 pagesClase 8 Lipogenesis 2014 PDFCarolina López ChinoNo ratings yet
- LIPIDOSDocument5 pagesLIPIDOSVC?No ratings yet
- Formación de Cuerpos Cetónicos (Cetogénesis)Document5 pagesFormación de Cuerpos Cetónicos (Cetogénesis)MilvetNo ratings yet
- 02A Regulación MetabólicaDocument40 pages02A Regulación MetabólicaJhordy MamaniNo ratings yet
- Biosíntesis de Carbohidratos - BIOQUIMICA II U2Document13 pagesBiosíntesis de Carbohidratos - BIOQUIMICA II U2Bernardo Morales0% (1)
- Mecanismo Intrínseco y ExtrínsecoDocument7 pagesMecanismo Intrínseco y ExtrínsecoAlvaro CordovaNo ratings yet
- Cadena Respiratoria y Fosforilación OxidativaDocument40 pagesCadena Respiratoria y Fosforilación OxidativaDaniela LeónNo ratings yet
- Especialización de La MembranaDocument2 pagesEspecialización de La MembranaMarisol100% (2)
- Taller de Bioquimica IncompletoDocument4 pagesTaller de Bioquimica Incompletolaura carmonaNo ratings yet
- Adhesion CelularDocument30 pagesAdhesion CelularBeto HdzNo ratings yet
- Discusion 3Document17 pagesDiscusion 3Milver EscalanteNo ratings yet
- RESPIRACIÓNDocument16 pagesRESPIRACIÓNRonald ChavezNo ratings yet
- Represion CatabolicaDocument25 pagesRepresion CatabolicaTarrillo Dávila Miguel AngelNo ratings yet
- Equipo 7 Práctica 4 Electroforesis RevisadoDocument14 pagesEquipo 7 Práctica 4 Electroforesis RevisadoSamuel SosaNo ratings yet
- Biosíntesis LípidosDocument17 pagesBiosíntesis LípidospopiraquelNo ratings yet
- Apuntes Tema 18Document8 pagesApuntes Tema 18gmolermaNo ratings yet
- Minimentaldef MMSE PDFDocument1 pageMinimentaldef MMSE PDFFernando Octavio Hernandez VallesNo ratings yet
- LKLLKKLLDocument25 pagesLKLLKKLLMarcelo SuarezNo ratings yet
- Sistema de EndomembranasDocument53 pagesSistema de EndomembranasMarcelo SuarezNo ratings yet
- Instructivo Tarea de Bioseguridad PDFDocument1 pageInstructivo Tarea de Bioseguridad PDFMarcelo SuarezNo ratings yet
- Instructivo Hidratantes - EnergizantesDocument1 pageInstructivo Hidratantes - EnergizantesMarcelo SuarezNo ratings yet
- Practica de Bioseguridad PDFDocument14 pagesPractica de Bioseguridad PDFMarcelo Suarez0% (1)
- Aplicación de La Metodología Científica en La Elaboración de Informes PDFDocument1 pageAplicación de La Metodología Científica en La Elaboración de Informes PDFMarcelo SuarezNo ratings yet
- Fisiologia - Cardiovascular IIIb - Ruidos Cardiacos y AuscultacionDocument1 pageFisiologia - Cardiovascular IIIb - Ruidos Cardiacos y AuscultacionCecilia PascoNo ratings yet
- ALCOHOLISMODocument1 pageALCOHOLISMOGran JaguarNo ratings yet
- Fisiologia - Cardiovascular I - Electrofisiologia Del CorazonDocument3 pagesFisiologia - Cardiovascular I - Electrofisiologia Del CorazonMarcelo SuarezNo ratings yet
- Fisiologia - Cardiovascular IIIa - Ciclo Cardiaco y Contraccion MuscularDocument2 pagesFisiologia - Cardiovascular IIIa - Ciclo Cardiaco y Contraccion MuscularAlan Chovanova VedderNo ratings yet
- Fisiologia - Cardiovascular II - ElectrocardiografiaDocument7 pagesFisiologia - Cardiovascular II - ElectrocardiografiaecleptosNo ratings yet
- Fisiologia - Cardiovascular VI - Fisiologia Del EndotelioDocument3 pagesFisiologia - Cardiovascular VI - Fisiologia Del EndotelioCinthia UrbinaNo ratings yet
- Fisiologia - Cardiovascular V - Regulacion de La Presion ArterialDocument5 pagesFisiologia - Cardiovascular V - Regulacion de La Presion ArterialAlan Chovanova VedderNo ratings yet
- Fisiologia - Cardiovascular IVa - Regulacion Del Volumen Minuto CardiacoDocument3 pagesFisiologia - Cardiovascular IVa - Regulacion Del Volumen Minuto CardiacoJamil Saavedra DiazNo ratings yet
- KKKKKDocument24 pagesKKKKKMarcelo SuarezNo ratings yet
- Fisiologia - Cardiovascular IVb - ExtritasDocument2 pagesFisiologia - Cardiovascular IVb - ExtritasFreddy Alexito BravoNo ratings yet
- Pruebas SerologicasDocument17 pagesPruebas SerologicasMarcelo SuarezNo ratings yet
- Grasas TransDocument5 pagesGrasas TransMarcelo SuarezNo ratings yet
- Ardor GástricoDocument3 pagesArdor GástricoMarcelo SuarezNo ratings yet
- ALCOHOLISMODocument1 pageALCOHOLISMOGran JaguarNo ratings yet
- Ciclo MenstrualDocument28 pagesCiclo MenstrualMarcelo SuarezNo ratings yet
- HC NacDocument4 pagesHC NacMarcelo SuarezNo ratings yet
- Ac. Araquidonico PGS Etc 1Document106 pagesAc. Araquidonico PGS Etc 1Marcelo SuarezNo ratings yet
- PCRDocument2 pagesPCRMarcelo SuarezNo ratings yet
- PatoDocument5 pagesPatoMarcelo SuarezNo ratings yet
- La Salud Es Un DerechoDocument2 pagesLa Salud Es Un DerechoMarcelo SuarezNo ratings yet
- Mapa ParlanteDocument29 pagesMapa ParlanteMarcelo Suarez0% (1)
- Huesos de La ManoDocument7 pagesHuesos de La ManoMarcelo Suarez100% (1)
- Documentación Sprint BootDocument6 pagesDocumentación Sprint BootGIULIANA MICHELLE HIDROBO HERRERANo ratings yet
- Indicadores en SaludDocument34 pagesIndicadores en SaludjorgenzapataNo ratings yet
- Un Caso AisladoDocument41 pagesUn Caso AisladoInmobiliaria Pilares Sociedad AnónimaNo ratings yet
- Epistologia 1,2,3Document18 pagesEpistologia 1,2,3ximenaNo ratings yet
- Declinacion Del Notario DominicanoDocument11 pagesDeclinacion Del Notario DominicanoJ.d. MatosNo ratings yet
- Calculo Hidraulico de Reservorio 13.5 m3Document4 pagesCalculo Hidraulico de Reservorio 13.5 m3José Eduardo Cruz GómezNo ratings yet
- Perfil de GrupoDocument42 pagesPerfil de GrupoLiliana Reyes ZamoraNo ratings yet
- Aritmetica 2Document48 pagesAritmetica 2Fabrizzio CayllahuaNo ratings yet
- Marx - Tesis Sobre FeuerbachDocument4 pagesMarx - Tesis Sobre FeuerbachNicolás AldunateNo ratings yet
- Estudios de Suelos - GuajiraDocument44 pagesEstudios de Suelos - GuajiraOscar Jose Carrillo CarreñoNo ratings yet
- Número de Documento NRF-030-PEMEX-2009 21 de JulioDocument104 pagesNúmero de Documento NRF-030-PEMEX-2009 21 de JulioluisgasNo ratings yet
- Guias Octavo Segundo Periodo PDocument7 pagesGuias Octavo Segundo Periodo PDiana Katherine MARTINEZ PAEZNo ratings yet
- Excel Básico Intermedio y AvanzadoDocument2 pagesExcel Básico Intermedio y Avanzadomario floresNo ratings yet
- Construcción de Terraplenes PDFDocument5 pagesConstrucción de Terraplenes PDFDaniel HuancaNo ratings yet
- Loza 2.3.1 PLT - Gs.se - Pets.ant 024 Servicio de Instalación de Oficina Contenedor Rev00.Document16 pagesLoza 2.3.1 PLT - Gs.se - Pets.ant 024 Servicio de Instalación de Oficina Contenedor Rev00.Clefa Cattleya LfNo ratings yet
- Monitoreo y Control Automático de Equipos IPDocument5 pagesMonitoreo y Control Automático de Equipos IPAnthony SahuanayNo ratings yet
- Una Propuesta de Comunicacion Por Juan Manuel Fernández MorenoDocument11 pagesUna Propuesta de Comunicacion Por Juan Manuel Fernández MorenoGermanGutierrez50% (2)
- Mantenimiento Centrado en Confiabilidad 1Document3 pagesMantenimiento Centrado en Confiabilidad 1Stalin OlmedoNo ratings yet
- Gcav U3 A3Document3 pagesGcav U3 A3Zoe Ron100% (1)
- Contrato MulticanalDocument7 pagesContrato MulticanalJuan Carlos GarciaNo ratings yet
- Crucigrama XDDocument1 pageCrucigrama XDNaira Vanessa Diaz HernandezNo ratings yet
- Catalogo NGK 2015 NovDocument47 pagesCatalogo NGK 2015 NovJohnny M. Pancha RamosNo ratings yet
- Actividad Virtual 3 TopografiaDocument4 pagesActividad Virtual 3 Topografiajaime100% (1)
- Trabajo Total de Proyecto de Innovacion Tecnologico FinalDocument9 pagesTrabajo Total de Proyecto de Innovacion Tecnologico FinalJuan Espinoza HernandezNo ratings yet
- Keto Rola CoDocument5 pagesKeto Rola CoMaricarmen Martínez100% (1)
- Catalogo - Desagues Canaletas CanalesDocument40 pagesCatalogo - Desagues Canaletas CanalesEsmeralda García FuentesNo ratings yet
- Actividad de Aprendizaje 1 Cyt 28-04 IiiexDocument7 pagesActividad de Aprendizaje 1 Cyt 28-04 IiiexLizet Sedano SotoNo ratings yet
- Guia Aritmetica 2Document1 pageGuia Aritmetica 2PABLO PARADANo ratings yet
- Chapter 7 - Enzymatic Quorum Quenching in Biofilms - 2019 - Quorum SensingDocument21 pagesChapter 7 - Enzymatic Quorum Quenching in Biofilms - 2019 - Quorum SensingFrancelia Quiñonez RuvalcabaNo ratings yet
- El Vale y Bonos Bancarios Donny ReyesDocument19 pagesEl Vale y Bonos Bancarios Donny ReyesRobertoReyesPerezNo ratings yet