You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Filipino FamilyDocument11 pagesThe Filipino FamilyTiger Knee97% (37)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Vedic Astrology OverviewDocument1 pageVedic Astrology Overviewhuman999100% (8)
- Bread Machine Sunbeam 5891Document44 pagesBread Machine Sunbeam 5891Tyler KirklandNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- German BasicDocument60 pagesGerman BasicchahirNo ratings yet
- FrankensteinDocument24 pagesFrankensteinLisa WardNo ratings yet
- ATLAS CYLINDER LUBRICATOR MANUALDocument36 pagesATLAS CYLINDER LUBRICATOR MANUALKaleb Z king webNo ratings yet
- Cbydp Draft SK BaracbacDocument13 pagesCbydp Draft SK BaracbacLikey PromiseNo ratings yet
- MATH 8 QUARTER 3 WEEK 1 & 2 MODULEDocument10 pagesMATH 8 QUARTER 3 WEEK 1 & 2 MODULECandy CastroNo ratings yet
- Analog To Digital Conversion (ADC)Document62 pagesAnalog To Digital Conversion (ADC)Asin PillaiNo ratings yet
- Openstack Deployment Ops Guide PDFDocument197 pagesOpenstack Deployment Ops Guide PDFBinank PatelNo ratings yet
- 2018 Cambridge Lower Second Progression Test Science Stage 8 QP Paper 2 - tcm143-430409Document16 pages2018 Cambridge Lower Second Progression Test Science Stage 8 QP Paper 2 - tcm143-430409AnisahNo ratings yet
- In 'T Zandt, Van Der Graaf, Heerschap - 2001 - Common Processing of in Vivo MR Spectra PDFDocument9 pagesIn 'T Zandt, Van Der Graaf, Heerschap - 2001 - Common Processing of in Vivo MR Spectra PDFjan5437No ratings yet
- Banks, Dinges - 2007 - Behavioral and Physiological Consequences of Sleep RestrictionDocument10 pagesBanks, Dinges - 2007 - Behavioral and Physiological Consequences of Sleep Restrictionjan5437No ratings yet
- Inoue Et Al. - 2017 - Glucocorticoid Receptor-Mediated Amygdalar Metaplasticity Underlies Adaptive Modulation of Fear Memory by Stress PDFDocument48 pagesInoue Et Al. - 2017 - Glucocorticoid Receptor-Mediated Amygdalar Metaplasticity Underlies Adaptive Modulation of Fear Memory by Stress PDFjan5437No ratings yet
- Hyvärinen, Oja - 2000 - Independent Component Analysis Algorithms and Applications PDFDocument20 pagesHyvärinen, Oja - 2000 - Independent Component Analysis Algorithms and Applications PDFjan5437No ratings yet
- NIH Public AccessDocument15 pagesNIH Public Accessjan5437No ratings yet
- Verhage Et Al. - 2000 - Synaptic Assembly of The Brain in The Absence of Neurotransmitter SecretionDocument7 pagesVerhage Et Al. - 2000 - Synaptic Assembly of The Brain in The Absence of Neurotransmitter Secretionjan5437No ratings yet
- Hunsaker, Tran, Kesner - 2009 - A Behavioral Analysis of The Role of CA3 and CA1 Subcortical Efferents During Classical Fear ConditioningDocument7 pagesHunsaker, Tran, Kesner - 2009 - A Behavioral Analysis of The Role of CA3 and CA1 Subcortical Efferents During Classical Fear Conditioningjan5437No ratings yet
- Hui Et Al. - 2004 - Memory Enhancement of Classical Fear Conditioning by Post-Training Injections of Corticosterone in RatsDocument8 pagesHui Et Al. - 2004 - Memory Enhancement of Classical Fear Conditioning by Post-Training Injections of Corticosterone in Ratsjan5437No ratings yet
- Hui Et Al. - 2006 - Posttraining Handling Facilitates Memory For Auditory-Cue Fear Conditioning in RatsDocument4 pagesHui Et Al. - 2006 - Posttraining Handling Facilitates Memory For Auditory-Cue Fear Conditioning in Ratsjan5437No ratings yet
- Vargas-Ashby, Pankhurst - 2007 - Effects of Feeding Enrichment On The Behaviour and Welfare of Captive Waldrapps (Northern Bald Ibis) (GeronDocument7 pagesVargas-Ashby, Pankhurst - 2007 - Effects of Feeding Enrichment On The Behaviour and Welfare of Captive Waldrapps (Northern Bald Ibis) (Geronjan5437100% (1)
- Verheyen Et Al. - 2019 - Motor Development in Children With Dravet SyndromeDocument8 pagesVerheyen Et Al. - 2019 - Motor Development in Children With Dravet Syndromejan5437No ratings yet
- Merck Sharp and Dohme Research Laboratories, Neuroscience Research Centre, Terlings Park, Harlow, Essex, United KingdomDocument13 pagesMerck Sharp and Dohme Research Laboratories, Neuroscience Research Centre, Terlings Park, Harlow, Essex, United Kingdomjan5437No ratings yet
- Hsu, Tomblin, Christiansen - 2014 - Impaired Statistical Learning of Non-Adjacent Dependencies in Adolescents With Specific Language ImpairmDocument10 pagesHsu, Tomblin, Christiansen - 2014 - Impaired Statistical Learning of Non-Adjacent Dependencies in Adolescents With Specific Language Impairmjan5437No ratings yet
- Vetere Et Al. - 2013 - Reactivating Fear Memory Under Propranolol Resets Pre-Trauma Levels of Dendritic Spines in Basolateral Amygdala But N PDFDocument9 pagesVetere Et Al. - 2013 - Reactivating Fear Memory Under Propranolol Resets Pre-Trauma Levels of Dendritic Spines in Basolateral Amygdala But N PDFjan5437No ratings yet
- Badcock Et Al. - 2012 - Co-Localisation of Abnormal Brain Structure and Function in Specific Language Impairment PDFDocument11 pagesBadcock Et Al. - 2012 - Co-Localisation of Abnormal Brain Structure and Function in Specific Language Impairment PDFjan5437No ratings yet
- Albouy Et Al. - 2013 - Daytime Sleep Enhances Consolidation of The Spatial But Not Motoric Representation of Motor Sequence MemoryDocument13 pagesAlbouy Et Al. - 2013 - Daytime Sleep Enhances Consolidation of The Spatial But Not Motoric Representation of Motor Sequence Memoryjan5437No ratings yet
- Merck Sharp and Dohme Research Laboratories, Neuroscience Research Centre, Terlings Park, Harlow, Essex, United KingdomDocument13 pagesMerck Sharp and Dohme Research Laboratories, Neuroscience Research Centre, Terlings Park, Harlow, Essex, United Kingdomjan5437No ratings yet
- Dong - 2015 - Proton MRS and MRSI of The Brain Without Water SuppressionDocument15 pagesDong - 2015 - Proton MRS and MRSI of The Brain Without Water Suppressionjan5437No ratings yet
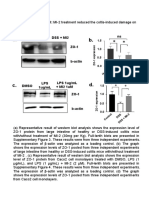
- Lee, Kim, Lee - 2018 - Treatment of Dextran Sulfate Sodium-Induced Colitis With Mucosa-Associated Lymphoid Tissue Lymphoma Translocation 1 I PDFDocument5 pagesLee, Kim, Lee - 2018 - Treatment of Dextran Sulfate Sodium-Induced Colitis With Mucosa-Associated Lymphoid Tissue Lymphoma Translocation 1 I PDFjan5437No ratings yet
- Shi Et Al. - 2016 - Beneficial Properties of ProbioticsDocument12 pagesShi Et Al. - 2016 - Beneficial Properties of Probioticsjan5437No ratings yet
- Batalla Et Al. - 2013 - Structural and Functional Imaging Studies in Chronic Cannabis Users A Systematic Review of Adolescent and Adult FindDocument18 pagesBatalla Et Al. - 2013 - Structural and Functional Imaging Studies in Chronic Cannabis Users A Systematic Review of Adolescent and Adult Findjan5437No ratings yet
- Andreychenko Et Al. - 2012 - Efficient Spectral Editing at 7 T GABA Detection With MEGA-sLASERDocument8 pagesAndreychenko Et Al. - 2012 - Efficient Spectral Editing at 7 T GABA Detection With MEGA-sLASERjan5437No ratings yet
- Banks, Dinges - 2007 - Behavioral and Physiological Consequences of Sleep RestrictionDocument10 pagesBanks, Dinges - 2007 - Behavioral and Physiological Consequences of Sleep Restrictionjan5437No ratings yet
- Arp Et Al. - 2014 - Mineralocorticoid Receptors Guide Spatial and Stimulus-Response Learning in MiceDocument11 pagesArp Et Al. - 2014 - Mineralocorticoid Receptors Guide Spatial and Stimulus-Response Learning in Micejan5437No ratings yet
- Anxiety Disorders and Gaba NeutransmiterDocument11 pagesAnxiety Disorders and Gaba Neutransmitercitra dewiNo ratings yet
- Arp Et Al. - 2014 - Mineralocorticoid Receptors Guide Spatial and Stimulus-Response Learning in MiceDocument11 pagesArp Et Al. - 2014 - Mineralocorticoid Receptors Guide Spatial and Stimulus-Response Learning in Micejan5437No ratings yet
- Bates Et Al. - 2007 - Supplemental Data of Elephants Classify Human Ethnic Groups by Odor and Garment ColorDocument36 pagesBates Et Al. - 2007 - Supplemental Data of Elephants Classify Human Ethnic Groups by Odor and Garment Colorjan5437No ratings yet
- Bates Et Al. - 2007 - Supplemental Data of Elephants Classify Human Ethnic Groups by Odor and Garment ColorDocument2 pagesBates Et Al. - 2007 - Supplemental Data of Elephants Classify Human Ethnic Groups by Odor and Garment Colorjan5437No ratings yet
- Bates Et Al. - 2007 - Supplemental Data of Elephants Classify Human Ethnic Groups by Odor and Garment ColorDocument2 pagesBates Et Al. - 2007 - Supplemental Data of Elephants Classify Human Ethnic Groups by Odor and Garment Colorjan5437No ratings yet
- Bates Et Al. - 2007 - Elephants Classify Human Ethnic Groups by Odor and Garment ColorDocument5 pagesBates Et Al. - 2007 - Elephants Classify Human Ethnic Groups by Odor and Garment Colorjan5437No ratings yet
- Lecturer No 1 - Transformer BasicDocument1 pageLecturer No 1 - Transformer Basiclvb123No ratings yet
- ICO Basic SyllabusDocument11 pagesICO Basic SyllabusRaúl Plasencia Salini100% (1)
- Format For Handout - Comparative Models of EducationDocument5 pagesFormat For Handout - Comparative Models of EducationAdrian AsiNo ratings yet
- The Teacher and The LearnerDocument23 pagesThe Teacher and The LearnerUnique Alegarbes Labra-SajolNo ratings yet
- Gizmotchy 3 Element Complete Assembly InstructionsDocument5 pagesGizmotchy 3 Element Complete Assembly InstructionsEuropaNo ratings yet
- Psyclone: Rigging & Tuning GuideDocument2 pagesPsyclone: Rigging & Tuning GuidelmagasNo ratings yet
- Learn R For Applied StatisticsDocument457 pagesLearn R For Applied StatisticsyasortyNo ratings yet
- Range of Muscle Work.Document54 pagesRange of Muscle Work.Salman KhanNo ratings yet
- Material Safety Data Sheet: Development Associates, IncDocument3 pagesMaterial Safety Data Sheet: Development Associates, IncDedi MulyadiNo ratings yet
- Ultra Slimpak G448-0002: Bridge Input Field Configurable IsolatorDocument4 pagesUltra Slimpak G448-0002: Bridge Input Field Configurable IsolatorVladimirNo ratings yet
- Limit Switch 1LX7001-J AZBILDocument8 pagesLimit Switch 1LX7001-J AZBILHoàng Sơn PhạmNo ratings yet
- Nec TutorialDocument5 pagesNec TutorialbheemasenaNo ratings yet
- Lesson Plan 160205 Jelly Snake PracticalDocument7 pagesLesson Plan 160205 Jelly Snake Practicalapi-309206763No ratings yet
- 1-2-Chemical Indicator of GeopolymerDocument4 pages1-2-Chemical Indicator of GeopolymerYazmin Alejandra Holguin CardonaNo ratings yet
- BH Tif03Document21 pagesBH Tif03Andres R. OlguinNo ratings yet
- RRLDocument4 pagesRRLTiltshifter ViNo ratings yet
- Hall Design For SpeechDocument1 pageHall Design For SpeechSarovar SaroNo ratings yet
- New Wordpad DocumentDocument6 pagesNew Wordpad DocumentJonelle D'melloNo ratings yet