You might also like
- Encapsulation and Controlled Release Technologies in Food SystemsFrom EverandEncapsulation and Controlled Release Technologies in Food SystemsDr Jamileh M. LakkisNo ratings yet
- Ramos-Suárez and Carreras (2014) - Use of Microalgae Residues For Biogas Production PDFDocument10 pagesRamos-Suárez and Carreras (2014) - Use of Microalgae Residues For Biogas Production PDFZouzo222No ratings yet
- Enzymes To Enhance ADSDocument4 pagesEnzymes To Enhance ADSDiana GarzónNo ratings yet
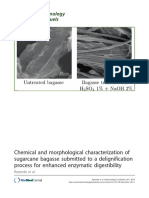
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- Biomass Pretreatment For Enhancement of Biogas Production: Tamilarasan Karuppiah and Vimala Ebenezer AzariahDocument22 pagesBiomass Pretreatment For Enhancement of Biogas Production: Tamilarasan Karuppiah and Vimala Ebenezer AzariahAimanNo ratings yet
- Drug Metabolism: CO NH H O Mineralisation Nutrients Amino Acids Toxins Polar Compounds ToxicokineticsDocument3 pagesDrug Metabolism: CO NH H O Mineralisation Nutrients Amino Acids Toxins Polar Compounds ToxicokineticssyarintaadeninaNo ratings yet
- Activated Sludge Response To Emulsified Lipid Loading: Steve E. HrudeyDocument13 pagesActivated Sludge Response To Emulsified Lipid Loading: Steve E. HrudeylucasNo ratings yet
- GRJ 2490Document6 pagesGRJ 2490Sri I.Balakrishna Assistant Professor (Sr.)No ratings yet
- Delignification, Bioconversion of Substrates and It's Affecting FactorsDocument32 pagesDelignification, Bioconversion of Substrates and It's Affecting FactorsHarshvardhan ZalaNo ratings yet
- Isolation of Bacterial Strain For Biodegradation of Fats, Oil and GreaseDocument6 pagesIsolation of Bacterial Strain For Biodegradation of Fats, Oil and GreaseraehanaeNo ratings yet
- International Journal of Biological MacromoleculesDocument13 pagesInternational Journal of Biological MacromoleculesMiguel Angel Espinace GarciasNo ratings yet
- MicrobialBioremediation PDFDocument20 pagesMicrobialBioremediation PDFMilenyRamosNo ratings yet
- Biomolecules: Lipases Immobilization For Effective Synthesis of Biodiesel Starting From Coffee Waste OilsDocument21 pagesBiomolecules: Lipases Immobilization For Effective Synthesis of Biodiesel Starting From Coffee Waste Oilsjuan diazNo ratings yet
- Biller2011 - Catalytic Hydrothermal Processing of Microalgae - DecompositionDocument8 pagesBiller2011 - Catalytic Hydrothermal Processing of Microalgae - Decompositionfarida_crNo ratings yet
- Technology For Treatment of Lipid-Rich Wastewater and Pipelines Clogged by Lipids Using Bacterial PreparationDocument9 pagesTechnology For Treatment of Lipid-Rich Wastewater and Pipelines Clogged by Lipids Using Bacterial PreparationAlacránAlemánNo ratings yet
- Bio TransformationDocument3 pagesBio Transformationmradu1No ratings yet
- Thesis M.SCDocument4 pagesThesis M.SCJameer InamdarNo ratings yet
- Suspended VS Attached Growth IJEE - Volume 7 - Issue 2 - Pages 114-123Document10 pagesSuspended VS Attached Growth IJEE - Volume 7 - Issue 2 - Pages 114-123Priyatno JuniantoNo ratings yet
- Kinetica Biodegradării Uleiului Petrolier de Către Un Consorțiu de Trei Izolate Protozoare (Aspidisca SP., Trachelophyllum Sp. Și Peranema SP.)Document7 pagesKinetica Biodegradării Uleiului Petrolier de Către Un Consorțiu de Trei Izolate Protozoare (Aspidisca SP., Trachelophyllum Sp. Și Peranema SP.)Madalina ElenaNo ratings yet
- Cheng 2017Document14 pagesCheng 2017Insanul KholikNo ratings yet
- Chemical Conversion of Biomass To Green ChemicalsDocument32 pagesChemical Conversion of Biomass To Green ChemicalsMaria Cecille Sarmiento GarciaNo ratings yet
- Monsal Enzymic Hydrolysis New Developments and Lessons LearntDocument23 pagesMonsal Enzymic Hydrolysis New Developments and Lessons LearntAnonymous MVHQ97KEoPNo ratings yet
- 21 Teka Et Al 2022 JAMB 22Document8 pages21 Teka Et Al 2022 JAMB 22Lekeah Pedro PehkatehNo ratings yet
- Biotechnology For Biofuels: Yield-Determining Factors in High-Solids Enzymatic Hydrolysis of LignocelluloseDocument10 pagesBiotechnology For Biofuels: Yield-Determining Factors in High-Solids Enzymatic Hydrolysis of LignocelluloseAlfonsius Dimas AdityoNo ratings yet
- Evaluation of Support Matrices For Immobilization of Anaerobic Consortia For e Cient Carbon Cycling in Waste RegenerationDocument10 pagesEvaluation of Support Matrices For Immobilization of Anaerobic Consortia For e Cient Carbon Cycling in Waste RegenerationGregorius BudiantoNo ratings yet
- Food Chemistry & Biochemistry (I) 1&2 Units FT 20Document24 pagesFood Chemistry & Biochemistry (I) 1&2 Units FT 20eruditeramanaNo ratings yet
- Polymers: Production of Low Molecular Weight Chitosan and Chitooligosaccharides (COS) : A ReviewDocument23 pagesPolymers: Production of Low Molecular Weight Chitosan and Chitooligosaccharides (COS) : A ReviewDenis Crispin IrazabalNo ratings yet
- GOOD Pretreatment BRTL18-19Document28 pagesGOOD Pretreatment BRTL18-19Ayush ModiNo ratings yet
- (ARTICULO) Caracterizacion de Lipasa Mucor Racemosus Con Aplicacion Potencial en El Tratamiento de La CelulitisDocument7 pages(ARTICULO) Caracterizacion de Lipasa Mucor Racemosus Con Aplicacion Potencial en El Tratamiento de La CelulitisAle EstradaNo ratings yet
- Ijmr 3 (1) 5-10Document6 pagesIjmr 3 (1) 5-10jeevalakshmanan29No ratings yet
- Anaerobic Digestion: Biology and BenefitsDocument10 pagesAnaerobic Digestion: Biology and Benefitswalidghoneim1970No ratings yet
- Principles of Anaerobic Digestio1Document18 pagesPrinciples of Anaerobic Digestio1ashe zinabNo ratings yet
- Basyaruddin 23 3 1Document7 pagesBasyaruddin 23 3 1NiegleNo ratings yet
- Tongco2020 Article EnhancementOfHydrolysisAndBiogDocument9 pagesTongco2020 Article EnhancementOfHydrolysisAndBiogJim TsikasNo ratings yet
- Optimization SpringerDocument10 pagesOptimization SpringerRachelle Wong Sue YeeNo ratings yet
- Gonzales 2017Document10 pagesGonzales 2017Thayna CandidoNo ratings yet
- 1 s2.0 S0306261912004266 Main.41Document6 pages1 s2.0 S0306261912004266 Main.41luuphuongNo ratings yet
- Advances in LipidDocument11 pagesAdvances in LipidAfifah N AhmedNo ratings yet
- Presented By:: Simerpreet Kaur Prachi Pandey Kumari Priya Shubham GuptaDocument12 pagesPresented By:: Simerpreet Kaur Prachi Pandey Kumari Priya Shubham Guptaalok sahuNo ratings yet
- LipozomiDocument12 pagesLipozomiDragota MihaiNo ratings yet
- Lipids Improving Oral BioavailabilityDocument23 pagesLipids Improving Oral BioavailabilityGary AgnewNo ratings yet
- Biostimulation: Microbial ConsortiumDocument4 pagesBiostimulation: Microbial ConsortiumDharmendra SinghNo ratings yet
- Life Cycle Analysis of Algae BiodieselDocument3 pagesLife Cycle Analysis of Algae BiodieselSiddhant SinghalNo ratings yet
- Improvement of Enzymatic SaccharificationDocument11 pagesImprovement of Enzymatic SaccharificationpepandradeNo ratings yet
- Pretreatment Methods 26.10.2022Document20 pagesPretreatment Methods 26.10.2022Bandita DashNo ratings yet
- Balasubramanian 2013Document8 pagesBalasubramanian 2013asadNo ratings yet
- Chapter 20.3-20.36 Revised - 6th Edition PDFDocument34 pagesChapter 20.3-20.36 Revised - 6th Edition PDFdzari6738No ratings yet
- Disposition and Crystallization of Saturated Fatty Acid in Mixed Micelles of Relevance To Lipid DigestionDocument7 pagesDisposition and Crystallization of Saturated Fatty Acid in Mixed Micelles of Relevance To Lipid DigestionRaul DavilaNo ratings yet
- Antioxidants 10 01587 v2Document19 pagesAntioxidants 10 01587 v2GILLIAN MERIDORNo ratings yet
- Using Dielectric Constant Measurement To Monitor Ethanol Fermentation and Anaerobic Co-Digestion of Lignocellulosic BiomassDocument20 pagesUsing Dielectric Constant Measurement To Monitor Ethanol Fermentation and Anaerobic Co-Digestion of Lignocellulosic Biomasszoltanjakoi.mkNo ratings yet
- International Biodeterioration & Biodegradation: Yongrui Pi, Long Meng, Mutai Bao, Peiyan Sun, Jinren LuDocument11 pagesInternational Biodeterioration & Biodegradation: Yongrui Pi, Long Meng, Mutai Bao, Peiyan Sun, Jinren LuAslam MuhammadNo ratings yet
- Impact of Seed Bioaugmentation On Thermophilic Anaerobic Digesters During Startup and Steady-StateDocument10 pagesImpact of Seed Bioaugmentation On Thermophilic Anaerobic Digesters During Startup and Steady-StateMario WhoeverNo ratings yet
- Sustainability 15 06832Document20 pagesSustainability 15 06832Piero Omar Obregón EscuderoNo ratings yet
- Dairy Industry-Anaerobic DigestorDocument16 pagesDairy Industry-Anaerobic Digestorneeraj sharmaNo ratings yet
- Bio DegrabilityDocument4 pagesBio DegrabilityMukesh LoharNo ratings yet
- Application of Bioemulsifiers in Soil Oil Bioremediation Processes. Future ProspectsDocument7 pagesApplication of Bioemulsifiers in Soil Oil Bioremediation Processes. Future ProspectsAndressa DecesaroNo ratings yet
- Separation and Purification TechnologyDocument7 pagesSeparation and Purification TechnologyIka SulistyaningtiyasNo ratings yet
- Ferment Able SugarDocument6 pagesFerment Able SugarXenia MenaNo ratings yet
- A Review of The Mechanism of Microbial Degradation of Petroleum PollutionDocument5 pagesA Review of The Mechanism of Microbial Degradation of Petroleum PollutionFIORELLA MARYCIELO CIPRIANO RAMOSNo ratings yet
- Chapter Ix. Single-Cell Protein and Ethanol Production: A. IntroductionDocument16 pagesChapter Ix. Single-Cell Protein and Ethanol Production: A. IntroductionattapharmacistNo ratings yet
- Apple Coleslaw - Chelsea's Messy ApronDocument3 pagesApple Coleslaw - Chelsea's Messy ApronJim TsikasNo ratings yet
- Lynmouth STW 2003Document2 pagesLynmouth STW 2003Jim TsikasNo ratings yet
- Wisa2002 P165Document9 pagesWisa2002 P165Jim TsikasNo ratings yet
- A La Carte Spring 2021Document1 pageA La Carte Spring 2021jatmehtaNo ratings yet
- Bacon Linguine Amatriciana 61162ecb068ea2167c5aeb21 3f6654ddDocument2 pagesBacon Linguine Amatriciana 61162ecb068ea2167c5aeb21 3f6654ddJim TsikasNo ratings yet
- Super Cheesy Oven-Baked Tomato Risotto: With Rocket SaladDocument2 pagesSuper Cheesy Oven-Baked Tomato Risotto: With Rocket SaladJim TsikasNo ratings yet
- OWASA Mason Farm Hydraulic and Treatment Capacity StudyDocument657 pagesOWASA Mason Farm Hydraulic and Treatment Capacity StudyJim TsikasNo ratings yet
- Lynmouth STW 2003Document2 pagesLynmouth STW 2003Jim TsikasNo ratings yet
- Hydrolysis Kinetics in Anaerobic Degradation of Particulate Organic Material: An Overview (J)Document14 pagesHydrolysis Kinetics in Anaerobic Degradation of Particulate Organic Material: An Overview (J)Jim TsikasNo ratings yet
- BGCprojectGTP EIS AppendixIDocument127 pagesBGCprojectGTP EIS AppendixIJim TsikasNo ratings yet
- Klargester Septic - Tanks Brochure - UK - Nov2016Document6 pagesKlargester Septic - Tanks Brochure - UK - Nov2016Jim TsikasNo ratings yet
- 0806A Claim WSX02 Sewage Treatment Works Capacity ProgrammeDocument183 pages0806A Claim WSX02 Sewage Treatment Works Capacity ProgrammeJim TsikasNo ratings yet
- Effects of Thermal Treatments On Five Different Waste Activated Sludge Samples Solubilisation, Physical Properties and Anaerobic DigestionDocument9 pagesEffects of Thermal Treatments On Five Different Waste Activated Sludge Samples Solubilisation, Physical Properties and Anaerobic DigestionJim TsikasNo ratings yet
- Hydrolytic Enzymes in Sewage Sludge Treatment: A Mini-ReviewDocument8 pagesHydrolytic Enzymes in Sewage Sludge Treatment: A Mini-ReviewJim TsikasNo ratings yet
- Install GuideDocument17 pagesInstall GuideJim TsikasNo ratings yet
- Pplication Ulletin: Lakos Saves Golf Course Dream House From Sand Trap NightmareDocument1 pagePplication Ulletin: Lakos Saves Golf Course Dream House From Sand Trap NightmareJim TsikasNo ratings yet
- Epanet and Development. A Progressive 44 Exercise WorkbookDocument198 pagesEpanet and Development. A Progressive 44 Exercise Workbookjcbobeda100% (2)
- Chemical Engineering Journal: Yonggang Xue, Huajie Liu, Sisi Chen, Norbert Dichtl, Xiaohu Dai, Ning LiDocument7 pagesChemical Engineering Journal: Yonggang Xue, Huajie Liu, Sisi Chen, Norbert Dichtl, Xiaohu Dai, Ning LiJim TsikasNo ratings yet
- Tongco2020 Article EnhancementOfHydrolysisAndBiogDocument9 pagesTongco2020 Article EnhancementOfHydrolysisAndBiogJim TsikasNo ratings yet
- Analysis of FOG Effect in Activated Sludge ProcessDocument11 pagesAnalysis of FOG Effect in Activated Sludge ProcessJim TsikasNo ratings yet
- ABB Softstarters PSEDocument13 pagesABB Softstarters PSEJim TsikasNo ratings yet
- Cyclone - SysCAD 2 DocumentationDocument13 pagesCyclone - SysCAD 2 DocumentationJim TsikasNo ratings yet
- Uq337846 OaDocument47 pagesUq337846 OaJim TsikasNo ratings yet
- Hydrostatic Test CertificateDocument2 pagesHydrostatic Test CertificateSulistyo Adi33% (3)
- 2013 Sept IHEDocument41 pages2013 Sept IHEJim TsikasNo ratings yet
- Dirtiness IndexDocument18 pagesDirtiness IndexJim TsikasNo ratings yet
- O-Ring Basic EN 2012 PDFDocument52 pagesO-Ring Basic EN 2012 PDFJim TsikasNo ratings yet
- O Ring GB en PDFDocument193 pagesO Ring GB en PDFJim TsikasNo ratings yet
- Veolia Containerised WTP BrochureDocument8 pagesVeolia Containerised WTP BrochureJim TsikasNo ratings yet
- 1418833938445Document12 pages1418833938445Jim TsikasNo ratings yet
- Cheetham Salt BrochureDocument8 pagesCheetham Salt BrochureSamscribdingNo ratings yet
- Vinyl Strip FlooringDocument2 pagesVinyl Strip Flooringjuli_radNo ratings yet
- In Vitro Anti-Inflammatory and Anti-Oxidant ActivitiesDocument11 pagesIn Vitro Anti-Inflammatory and Anti-Oxidant ActivitiesnimadelionNo ratings yet
- Joint Universities Preliminary Examinations Board 2015 Examinations Chemistry: Sci-J153 Multiple Choice Questions: Answer All QuestionsDocument11 pagesJoint Universities Preliminary Examinations Board 2015 Examinations Chemistry: Sci-J153 Multiple Choice Questions: Answer All QuestionsDeborahNo ratings yet
- AlSi12Fe MS, Props Under VacuumDocument14 pagesAlSi12Fe MS, Props Under VacuumRishabh MenonNo ratings yet
- PPG Cs 743: PPG CS 743 Is A Copolymer Polyol Especially Designed For The Production ofDocument2 pagesPPG Cs 743: PPG CS 743 Is A Copolymer Polyol Especially Designed For The Production ofVaittianathan MahavapillaiNo ratings yet
- Citrus Bioflavanoids 60% (FEMAS) : Cambridge CommoditiesDocument16 pagesCitrus Bioflavanoids 60% (FEMAS) : Cambridge Commoditiesibad muhammedNo ratings yet
- ANSYS Mechanical APDL Fracture Analysis GuideDocument134 pagesANSYS Mechanical APDL Fracture Analysis Guidearulrajasi0% (1)
- Quality Control Answer Key PINK PACOPDocument40 pagesQuality Control Answer Key PINK PACOPKharel Ezra de Guzman67% (3)
- Studies On Chemical Resistance of Low Water/cement Ratio ConcretesDocument10 pagesStudies On Chemical Resistance of Low Water/cement Ratio ConcretesMaziar MehravarNo ratings yet
- Solder Re Flow Guide For Surface Mount DevicesDocument13 pagesSolder Re Flow Guide For Surface Mount DevicesWandyy GonçalvesNo ratings yet
- The Delta RatioDocument2 pagesThe Delta RatioFirstglobalsupercopNo ratings yet
- Greentech - Isolatek SBK-113 TDSDocument2 pagesGreentech - Isolatek SBK-113 TDSRANo ratings yet
- Types of Chemical ReactionsDocument35 pagesTypes of Chemical ReactionsJemina R. B. EspedillonNo ratings yet
- 1989 90 PublicationDocument98 pages1989 90 PublicationCassanova SmileNo ratings yet
- DesalinizaciónDocument48 pagesDesalinizaciónLoren AlayoNo ratings yet
- Antimicrobial Finish in TextilesDocument7 pagesAntimicrobial Finish in Textilesjohn_dominic_4No ratings yet
- Soal Rancang HeDocument2 pagesSoal Rancang HeDimas0% (1)
- Hydrocarbon and Alkyl Halide-1Document10 pagesHydrocarbon and Alkyl Halide-1Aarya Vardhan ShandilyaNo ratings yet
- CV - Anil K Shukla (Nih, Usa)Document6 pagesCV - Anil K Shukla (Nih, Usa)Mehedi HossainNo ratings yet
- Soal Soal Kimia Kelas 10Document4 pagesSoal Soal Kimia Kelas 10rohmatul aziziNo ratings yet
- MCQs All With KeysDocument47 pagesMCQs All With KeysSana UllahNo ratings yet
- TanninsDocument73 pagesTanninsOmkar JoshiNo ratings yet
- Chock FastDocument25 pagesChock FastBrajko NikolaNo ratings yet
- Solid Fuel SeriesDocument15 pagesSolid Fuel SeriesCesar ValenciaNo ratings yet
- Ugiweb 52492Document5 pagesUgiweb 52492badrNo ratings yet
- G-protein-Coupled ReceptorsDocument24 pagesG-protein-Coupled ReceptorsNaimi Amalia hatimahNo ratings yet
- HITEC University Electrical Engineering Submitted byDocument6 pagesHITEC University Electrical Engineering Submitted bysana majidNo ratings yet
- The Requisites Nuclear Medicine 4th (Elsevier, 2014)Document456 pagesThe Requisites Nuclear Medicine 4th (Elsevier, 2014)Sabrina Moraru100% (4)