You might also like
- Musculoskeletal Anatomy and Physiology 4Document67 pagesMusculoskeletal Anatomy and Physiology 4Dennis Nabor Muñoz, RN,RM100% (1)
- Energy Transformation PPTDocument21 pagesEnergy Transformation PPTmansfieldchen100% (1)
- Musculoskeletal Anatomy and Physiology 5Document36 pagesMusculoskeletal Anatomy and Physiology 5Dennis Nabor Muñoz, RN,RMNo ratings yet
- Midwifery Practice TestDocument51 pagesMidwifery Practice TestDennis Nabor Muñoz, RN,RM100% (1)
- Chapter22 LectureDocument45 pagesChapter22 Lecturecee_prasetyo100% (1)
- Foundations in Microbiology: Miscellaneous Bacterial Agents of Disease TalaroDocument46 pagesFoundations in Microbiology: Miscellaneous Bacterial Agents of Disease TalaroOdurNo ratings yet
- Foundations in Microbiology: The Cocci of Medical Importance TalaroDocument71 pagesFoundations in Microbiology: The Cocci of Medical Importance TalaroOdurNo ratings yet
- Foundations in Microbiology: The Gram-Positive Bacilli of Medical Importance TalaroDocument65 pagesFoundations in Microbiology: The Gram-Positive Bacilli of Medical Importance TalaroOdurNo ratings yet
- 7 Environmental PrinciplesDocument26 pages7 Environmental PrinciplesDaxNo ratings yet
- DeforestationDocument2 pagesDeforestationNabila NazmaNo ratings yet
- Design Inputs for 550 CMD SBR Wastewater Treatment PlantDocument5 pagesDesign Inputs for 550 CMD SBR Wastewater Treatment PlantHemant KaleNo ratings yet
- Foundations in Microbiology: Microbe-Human Interactions: Infection and Disease TalaroDocument46 pagesFoundations in Microbiology: Microbe-Human Interactions: Infection and Disease TalaroOdurNo ratings yet
- The Gram-Negative Bacilli of Medical ImportanceDocument54 pagesThe Gram-Negative Bacilli of Medical ImportanceAna-Maria Nicolae100% (1)
- Foundations in Microbiology: Drugs, Microbes, Host - The Elements of Chemotherapy TalaroDocument66 pagesFoundations in Microbiology: Drugs, Microbes, Host - The Elements of Chemotherapy TalaroOdurNo ratings yet
- Foundations in Microbiology: Nonspecific Host Defenses TalaroDocument35 pagesFoundations in Microbiology: Nonspecific Host Defenses TalaroOdurNo ratings yet
- Colston Et Al-2016-Molecular Ecology2222Document25 pagesColston Et Al-2016-Molecular Ecology2222Diana CalderaNo ratings yet
- American Arachnological SocietyDocument8 pagesAmerican Arachnological SocietyEvan SadnessNo ratings yet
- Osório Et Al., 2018 - Periphytic Cosmarium (Zygnematophyceae, Desmidiaceae) ...Document11 pagesOsório Et Al., 2018 - Periphytic Cosmarium (Zygnematophyceae, Desmidiaceae) ...Ubirajara FernandesNo ratings yet
- Nematode Community Structure As A BioindDocument5 pagesNematode Community Structure As A BioindAldy AsraNo ratings yet
- Extinctions of Interactions Quantifying A Dung BeeDocument15 pagesExtinctions of Interactions Quantifying A Dung BeeTossyNo ratings yet
- Mycorrhizal NetworksDocument22 pagesMycorrhizal NetworkscharlytogfNo ratings yet
- Akoueson 2020Document10 pagesAkoueson 2020jhellianeteves14No ratings yet
- Molecular Ecology - 2013 - Delsuc - Convergence of Gut Microbiomes in Myrmecophagous MammalsDocument17 pagesMolecular Ecology - 2013 - Delsuc - Convergence of Gut Microbiomes in Myrmecophagous MammalsleandrasapucaiasNo ratings yet
- Effects of Insecticide on Amphibians at Different Densities and with PredationDocument11 pagesEffects of Insecticide on Amphibians at Different Densities and with PredationlektulariusNo ratings yet
- Insect Food WebsDocument43 pagesInsect Food WebsmaricarmenherrerafuentesNo ratings yet
- Deleting Species From Model Food WebsDocument14 pagesDeleting Species From Model Food WebsChuchitoLindoNo ratings yet
- Correlations Between Lizard Cranial Shape and Diet: A Quantitative, Phylogenetically Informed AnalysisDocument34 pagesCorrelations Between Lizard Cranial Shape and Diet: A Quantitative, Phylogenetically Informed Analysisalexandra gonzalez castilloNo ratings yet
- Agriculture, Ecosystems and Environment: A B C A A D 1 A 1Document8 pagesAgriculture, Ecosystems and Environment: A B C A A D 1 A 1Evan SadnessNo ratings yet
- M. Aldana (2020)Document8 pagesM. Aldana (2020)steeven.j.alejandroNo ratings yet
- Invertebrate - An OverviewDocument13 pagesInvertebrate - An OverviewJenna PretalNo ratings yet
- 1 s2.0 S0031018221004211 MainDocument16 pages1 s2.0 S0031018221004211 MainAtiqah AuliaNo ratings yet
- 2020 ReferenceDocument12 pages2020 ReferenceRayne LoefflerNo ratings yet
- Ferguson Joly 2002Document9 pagesFerguson Joly 2002Wendy BautistaNo ratings yet
- Conservation Ecology of Bees Populations, Species and CommunitiesDocument26 pagesConservation Ecology of Bees Populations, Species and CommunitiesanradeljaNo ratings yet
- Uwadiae 2011Document19 pagesUwadiae 2011davidNo ratings yet
- Williams2010 PDFDocument12 pagesWilliams2010 PDFFernando Forster FurquimNo ratings yet
- 2 Tiselius and Moller JPR 2017 Community Cascades Caused by CtenophoreDocument9 pages2 Tiselius and Moller JPR 2017 Community Cascades Caused by CtenophoreRubyHudkinsNo ratings yet
- Burkepile Oikos 2013Document7 pagesBurkepile Oikos 2013Mark AmbuangNo ratings yet
- Functional Diversity of Crustacean Zooplankton Communities Towards A Trait-Based Classification 2007 BARNETT Marcado PdfviewDocument18 pagesFunctional Diversity of Crustacean Zooplankton Communities Towards A Trait-Based Classification 2007 BARNETT Marcado PdfviewrgelabertNo ratings yet
- Small Herbivores With Big Impacts: Tundra Voles (Microtus Oeconomus) Alter Post-Fire Ecosystem DynamicsDocument13 pagesSmall Herbivores With Big Impacts: Tundra Voles (Microtus Oeconomus) Alter Post-Fire Ecosystem Dynamicsapi-670001661No ratings yet
- Carnivore coexistence facilitated by spatial, dietary and behavioural partitioningDocument16 pagesCarnivore coexistence facilitated by spatial, dietary and behavioural partitioningИванАнато́лийФранкоNo ratings yet
- Journal Pre-Proof: Science of The Total EnvironmentDocument34 pagesJournal Pre-Proof: Science of The Total EnvironmentChrisYapNo ratings yet
- Grupos FuncionalesDocument14 pagesGrupos FuncionalesClara Isabel DelgadoNo ratings yet
- Kamath et al 2018 Collective behavior and colony persistence of social spiders depends on their enviroDocument9 pagesKamath et al 2018 Collective behavior and colony persistence of social spiders depends on their envirobiomontsehdzglzNo ratings yet
- Gomes Dos Santos Et Al - 2019 - TIAEDocument22 pagesGomes Dos Santos Et Al - 2019 - TIAESIXTO GUTIERREZNo ratings yet
- Symbiotic InteractionsDocument18 pagesSymbiotic InteractionsJotha-OriginNo ratings yet
- Herbivora MamaliaDocument12 pagesHerbivora MamaliaselviNo ratings yet
- Molecular Ecology - 2011 - ZINGER - Two Decades of Describing The Unseen Majority of Aquatic Microbial DiversityDocument19 pagesMolecular Ecology - 2011 - ZINGER - Two Decades of Describing The Unseen Majority of Aquatic Microbial DiversityMihu MălinaNo ratings yet
- Ecologia Trofica Del E. MultilocularisDocument10 pagesEcologia Trofica Del E. MultiloculariscamiloesferNo ratings yet
- Role of Aquatic Plants in Structuring Periphytic MicroalgaeDocument8 pagesRole of Aquatic Plants in Structuring Periphytic MicroalgaeUbirajara FernandesNo ratings yet
- Estuarine, Coastal and Shelf Science: Christina Buelow, Marcus SheavesDocument11 pagesEstuarine, Coastal and Shelf Science: Christina Buelow, Marcus SheavesMaría Alejandra Parrado-VargasNo ratings yet
- 11-35 - 2 Trama (175-89)Document15 pages11-35 - 2 Trama (175-89)flortramaNo ratings yet
- Nesting Strategies and Disease Risk in Necrophagous BeetlesDocument15 pagesNesting Strategies and Disease Risk in Necrophagous BeetlesEduardo ValentimNo ratings yet
- Viruses and Nutritent Cycles in The Sea, Wilhelm & Suttle, 1999Document8 pagesViruses and Nutritent Cycles in The Sea, Wilhelm & Suttle, 1999deryhermawanNo ratings yet
- Evaluating Potential Artifacts of Tethering Techniques To Estimate Predation On Sea UrchinsDocument6 pagesEvaluating Potential Artifacts of Tethering Techniques To Estimate Predation On Sea UrchinsJulian RodriguezNo ratings yet
- 1 s2.0 S0301479723020492 MainDocument9 pages1 s2.0 S0301479723020492 Mainabdulbaess keyhaniNo ratings yet
- Boyero Et Al. 2020. Microplastics Impair Amphibian Survival, Body Condition and FunctionDocument7 pagesBoyero Et Al. 2020. Microplastics Impair Amphibian Survival, Body Condition and Functionprevencionista servimanefNo ratings yet
- Freshwater Biology - 2010 - PALMER - River Restoration Habitat Heterogeneity and Biodiversity A Failure of Theory orDocument18 pagesFreshwater Biology - 2010 - PALMER - River Restoration Habitat Heterogeneity and Biodiversity A Failure of Theory oraraNo ratings yet
- Chemosphere: Carolina Valdespino, Vinicio J. SosaDocument10 pagesChemosphere: Carolina Valdespino, Vinicio J. SosaLauraCristinaPachecoNo ratings yet
- Beyond_bacteria_reconstructing_microorganism_conneDocument11 pagesBeyond_bacteria_reconstructing_microorganism_conneElías Mindreau GanozaNo ratings yet
- 爬行动物和益生菌Document22 pages爬行动物和益生菌Xiaoqiang KeNo ratings yet
- Linking Species Richness, Biodiversity and Ecosystem Function in Soil SystemsDocument19 pagesLinking Species Richness, Biodiversity and Ecosystem Function in Soil SystemsroseNo ratings yet
- The Relevance of Ecological Pyramids in Community AssemblagesDocument9 pagesThe Relevance of Ecological Pyramids in Community AssemblagesAnonymous enxlMjs1No ratings yet
- Gill Et Al-2014-Functional EcologyDocument13 pagesGill Et Al-2014-Functional EcologyAlexandria TrahanNo ratings yet
- Molecular Ecology - 2018 - Bhattacharyya - A Diet Rich in C3 Plants Reveals The Sensitivity of An Alpine Mammal To ClimateDocument16 pagesMolecular Ecology - 2018 - Bhattacharyya - A Diet Rich in C3 Plants Reveals The Sensitivity of An Alpine Mammal To Climateraja sekhara reddy ravuriNo ratings yet
- Global Diversity and Distribution of Arbuscular Mycorrhizal FungiDocument10 pagesGlobal Diversity and Distribution of Arbuscular Mycorrhizal FungiErika MtzNo ratings yet
- Martinez de La Escalera Et Al 2016 Dynamics Toxic GenotypesDocument11 pagesMartinez de La Escalera Et Al 2016 Dynamics Toxic GenotypescarlosNo ratings yet
- Module 6 Trends No ActivityDocument6 pagesModule 6 Trends No Activitygirlie jimenezNo ratings yet
- Baker 2019Document12 pagesBaker 2019Sara Abigail Mateo OzorioNo ratings yet
- Uribeetal 2015 ADocument13 pagesUribeetal 2015 AMaritza Fajardo EscobarNo ratings yet
- A Morphological Classification CapturingDocument14 pagesA Morphological Classification CapturingfrfunkNo ratings yet
- 2012 Microbes ReportDocument17 pages2012 Microbes ReportOscar SerranoNo ratings yet
- Chapter 6Document28 pagesChapter 6AgnesNo ratings yet
- Seminar (Entomolgy 2)Document3 pagesSeminar (Entomolgy 2)Mehram JuttNo ratings yet
- Improve Your Medication Management Processes: Advisory ServicesDocument2 pagesImprove Your Medication Management Processes: Advisory ServicesSimarpreet KaurNo ratings yet
- Care of Clients With Problems Related To The Musculoskeletal SystemDocument46 pagesCare of Clients With Problems Related To The Musculoskeletal SystemMichelle Joy Mallillin MadriaNo ratings yet
- Musculoskeletal Anatomy and Physiology 2Document18 pagesMusculoskeletal Anatomy and Physiology 2Dennis Nabor Muñoz, RN,RMNo ratings yet
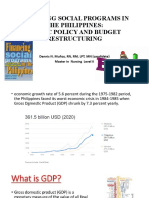
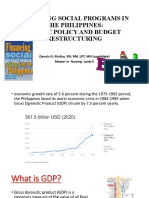
- Financing Social Programs in The Philippines: Public Policy and Budget RestructuringDocument78 pagesFinancing Social Programs in The Philippines: Public Policy and Budget RestructuringDennis Nabor Muñoz, RN,RMNo ratings yet
- Musculoskeletal Anatomy and Physiology 8Document41 pagesMusculoskeletal Anatomy and Physiology 8Dennis Nabor Muñoz, RN,RMNo ratings yet
- Financial Education Report 2021 Part 1Document28 pagesFinancial Education Report 2021 Part 1Dennis Nabor Muñoz, RN,RMNo ratings yet
- Musculoskeletal Anatomy and Physiology 7Document35 pagesMusculoskeletal Anatomy and Physiology 7Dennis Nabor Muñoz, RN,RMNo ratings yet
- Covid 19: Dennis N. Muñoz, RN, RM, LPTDocument103 pagesCovid 19: Dennis N. Muñoz, RN, RM, LPTDennis Nabor Muñoz, RN,RMNo ratings yet
- Musculoskeletal Anatomy and Physiology 2Document18 pagesMusculoskeletal Anatomy and Physiology 2Dennis Nabor Muñoz, RN,RMNo ratings yet
- Financial Education Report 2021 Part 2Document51 pagesFinancial Education Report 2021 Part 2Dennis Nabor Muñoz, RN,RMNo ratings yet
- Financing Social Programs in The Philippines: Public Policy and Budget RestructuringDocument78 pagesFinancing Social Programs in The Philippines: Public Policy and Budget RestructuringDennis Nabor Muñoz, RN,RMNo ratings yet
- Successful StoriesDocument5 pagesSuccessful StoriesDennis Nabor Muñoz, RN,RMNo ratings yet
- Financial Education Report 2021 Part 1.1Document14 pagesFinancial Education Report 2021 Part 1.1Dennis Nabor Muñoz, RN,RMNo ratings yet
- Foundations in Microbiology: Introduction To Viruses That Infect Humans: The DNA Viruses TalaroDocument50 pagesFoundations in Microbiology: Introduction To Viruses That Infect Humans: The DNA Viruses TalaroDennis Nabor Muñoz, RN,RMNo ratings yet
- Financial Education Report 2021 Part 1.2Document15 pagesFinancial Education Report 2021 Part 1.2Dennis Nabor Muñoz, RN,RMNo ratings yet
- Chapter23 LectureDocument74 pagesChapter23 Lecture7104No ratings yet
- Cell Surface MarkersDocument63 pagesCell Surface MarkersPallavi GhoshNo ratings yet
- Foundations in Microbiology: Diagnosing Infections TalaroDocument23 pagesFoundations in Microbiology: Diagnosing Infections TalaroOdurNo ratings yet
- Foundations in Microbiology: Disorders in Immunity TalaroDocument43 pagesFoundations in Microbiology: Disorders in Immunity TalaroOdurNo ratings yet
- Document 60Document2 pagesDocument 60Terrence LeeNo ratings yet
- Food Chain SimulationDocument5 pagesFood Chain SimulationJose CabezasNo ratings yet
- The Impact of Human Activities On Coral ReefsDocument19 pagesThe Impact of Human Activities On Coral ReefsmoniqueNo ratings yet
- Soal Uas Bahasa Inggris Kelas 11 Semester 1Document3 pagesSoal Uas Bahasa Inggris Kelas 11 Semester 1jumiospNo ratings yet
- Diversity and Classification of Indian Mangroves-Mandal - 2008 PDFDocument16 pagesDiversity and Classification of Indian Mangroves-Mandal - 2008 PDFBhatt SwetaNo ratings yet
- Forest AryDocument4 pagesForest ArySushant MoreNo ratings yet
- Ecoscenario EvergladesnpDocument17 pagesEcoscenario Evergladesnpapi-251354796No ratings yet
- Eflows BrochureDocument4 pagesEflows BrochureWanga SailiNo ratings yet
- 2011 Rodriguez-Olarte Et Al Fish Status Abrae VenezuelaDocument13 pages2011 Rodriguez-Olarte Et Al Fish Status Abrae VenezuelaDouglas Rodriguez OlarteNo ratings yet
- Ecosystems and Environmental IssuesDocument33 pagesEcosystems and Environmental IssuesAlfred Melvin Soliva100% (1)
- Unit 7 Ecology Study GuideDocument21 pagesUnit 7 Ecology Study Guidekylev0% (1)
- Ecological Engineering For Pest ManagementDocument9 pagesEcological Engineering For Pest ManagementAnggia AzizahNo ratings yet
- Mangrove Thesis 1st DraftDocument47 pagesMangrove Thesis 1st DraftMarie Dudan83% (6)
- Restoration Ecology and Conservation Biology (Young, 2000) PDFDocument11 pagesRestoration Ecology and Conservation Biology (Young, 2000) PDFJUANCHO_VANo ratings yet
- Types of EcosystemsDocument6 pagesTypes of EcosystemsPranavNo ratings yet
- Slide 1: Project Tiger Is A Wildlife Conservation MovementDocument9 pagesSlide 1: Project Tiger Is A Wildlife Conservation MovementPooja BkNo ratings yet
- ECOSYSTEM Environmental ScienceDocument6 pagesECOSYSTEM Environmental ScienceDonna MelgarNo ratings yet
- RMYA-ITEM-ANALYSIS-Gr4-6 MAPEHDocument10 pagesRMYA-ITEM-ANALYSIS-Gr4-6 MAPEHYani De SilvaNo ratings yet
- High and Low Biodiversity: ScienceDocument8 pagesHigh and Low Biodiversity: Scienceabegyll lolit micabaloNo ratings yet
- Community EcologyDocument4 pagesCommunity EcologyBiju ThomasNo ratings yet
- Mcqco 2Document9 pagesMcqco 2yayihih611No ratings yet
- Review On Status Potentials Threats and Challenges of The Fish Biodiversity of West BengalDocument10 pagesReview On Status Potentials Threats and Challenges of The Fish Biodiversity of West BengalDR. BIJAY KALI MAHAPATRANo ratings yet
- Evaluation of Water Equality of Major Rivers in Palawan Using Physico-Chemical Parameters and Water Quality IndexDocument13 pagesEvaluation of Water Equality of Major Rivers in Palawan Using Physico-Chemical Parameters and Water Quality IndexMark Ian EspirituNo ratings yet
- Jurnal Pengaruh Epifit Pada TemperaturDocument11 pagesJurnal Pengaruh Epifit Pada Temperatursosmed keluargaNo ratings yet
- Biology - Freeman 3E - Ecology Chapter 50 QuizDocument8 pagesBiology - Freeman 3E - Ecology Chapter 50 QuizEtoileCamelliaNo ratings yet
- Food Related Interactions: Grade 6Document14 pagesFood Related Interactions: Grade 6primalNo ratings yet