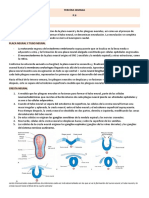
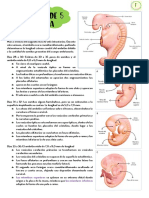
You might also like
- Laboratorio 4Document5 pagesLaboratorio 4nohemy.arguetaaNo ratings yet
- Proceso de formación de las capas germinales y estructuras embrionarias durante la gastrulaciónDocument4 pagesProceso de formación de las capas germinales y estructuras embrionarias durante la gastrulaciónMARIA FERNANDA RAZO GARCIANo ratings yet
- Desarrollo embrionario: las fases claveDocument36 pagesDesarrollo embrionario: las fases claveAnonymous 1Bpw6EK100% (1)
- EmbriologiaDocument5 pagesEmbriologiaFrank ReyesNo ratings yet
- Embriología Del Sistema Nervioso Central PDFDocument3 pagesEmbriología Del Sistema Nervioso Central PDFFrancis RamirezNo ratings yet
- NEURULACIÓN Juev. 10 de Dic.Document13 pagesNEURULACIÓN Juev. 10 de Dic.pablo cardonaNo ratings yet
- Periodo EmbrionarioDocument5 pagesPeriodo EmbrionarioFausty MaristaNo ratings yet
- Discigerminativo TrilaminarDocument23 pagesDiscigerminativo Trilaminargiuseppeperoni PichardoNo ratings yet
- De La Tercera A La Octava Semana..Document6 pagesDe La Tercera A La Octava Semana..Denilson Mena Tito ZabalaNo ratings yet
- Tema 7 NeuroanatoDocument10 pagesTema 7 NeuroanatoMaria Vara EnguitaNo ratings yet
- Desarrollo SNCDocument23 pagesDesarrollo SNCFelipe CantunNo ratings yet
- Embriologia Del Sistema NerviosoDocument72 pagesEmbriologia Del Sistema NerviosoburricorneoNo ratings yet
- Tercera Semana Del DesarrolloDocument6 pagesTercera Semana Del DesarrolloRaúl Gerardo De Lara GarciaNo ratings yet
- Desarrollo Embrionario PDFDocument11 pagesDesarrollo Embrionario PDFMoises Skazas Neuronas100% (1)
- De La Tercera A La Octava Semana Embrionaria y Hematopoyesis Fetal 327600 Downloable 519173 PDFDocument19 pagesDe La Tercera A La Octava Semana Embrionaria y Hematopoyesis Fetal 327600 Downloable 519173 PDFJeffrey LoayzaNo ratings yet
- Segunda Semana EmbrioDocument4 pagesSegunda Semana EmbrioMili GraNo ratings yet
- embrio de SNDocument9 pagesembrio de SNMalena AncillottiNo ratings yet
- Embriologia Disertacion-1Document31 pagesEmbriologia Disertacion-1Mamani M-aNo ratings yet
- Tema 5 Ontogenia Del Sistema NerviosoDocument10 pagesTema 5 Ontogenia Del Sistema NerviosoSandra GarcíaNo ratings yet
- Embriologia Del Sistema NerviosoDocument40 pagesEmbriologia Del Sistema NerviosoMartha Ligia Teherán HNo ratings yet
- 06 - Periodo EmbrionarioDocument49 pages06 - Periodo Embrionariolovers of everytingNo ratings yet
- 4dela3a8semanadeldesarrollo-Document77 pages4dela3a8semanadeldesarrollo-Andrea SotoNo ratings yet
- Desarrollo del Sistema Nervioso CentralDocument5 pagesDesarrollo del Sistema Nervioso Centraldaniela roldan pinoNo ratings yet
- 12 Capas GerminativasDocument22 pages12 Capas Germinativasgiuseppeperoni PichardoNo ratings yet
- EMBRIOLOGIA - Docx ExamenDocument3 pagesEMBRIOLOGIA - Docx Examenjose luis burgaNo ratings yet
- Capítulo 7, de La Tercera A La Octava SemanaDocument42 pagesCapítulo 7, de La Tercera A La Octava Semanajenny cruzNo ratings yet
- 4ta Semana Desarrollo EmbrionarioDocument27 pages4ta Semana Desarrollo EmbrionarioGeisy Mileidy ZavalaNo ratings yet
- Embriología de Moore: Tercera SemanaDocument2 pagesEmbriología de Moore: Tercera SemanaKaryme PérezNo ratings yet
- Ontogenia ResumenDocument17 pagesOntogenia ResumenEldy M. HenriquezNo ratings yet
- Cuadro Sinoptico 3a y 4a Semana de DesarrolloDocument3 pagesCuadro Sinoptico 3a y 4a Semana de DesarrolloReyes FcoNo ratings yet
- Gastrulacion y NeurulacionDocument6 pagesGastrulacion y NeurulacionVale MNo ratings yet
- Desarrollo Embriológico Del Sistema NerviosoDocument8 pagesDesarrollo Embriológico Del Sistema NerviosoErika Ruales100% (1)
- Capítulo 4. Tercera Semana Del Desarrollo HumanoDocument7 pagesCapítulo 4. Tercera Semana Del Desarrollo HumanoBarajas Gutiérrez MonserratNo ratings yet
- Periodo Embrionario: La etapa de formación de órganosDocument54 pagesPeriodo Embrionario: La etapa de formación de órganosFrida Ortega100% (1)
- Embriología Del SNCDocument5 pagesEmbriología Del SNCManuel Alejandro Parada100% (1)
- Resumen de Los 9 CapitulosDocument36 pagesResumen de Los 9 CapitulosSebastian Pinto MirandaNo ratings yet
- Embriología: Desarrollo de la 3ra semanaDocument18 pagesEmbriología: Desarrollo de la 3ra semanaFernanda Valeria Gutiérrez LeónNo ratings yet
- 5º La Gastrulación - Disco GerminativoDocument7 pages5º La Gastrulación - Disco GerminativoaleNo ratings yet
- Periodo SomiticoDocument45 pagesPeriodo SomiticoClaudio Skacion100% (1)
- Semana 3 Parte 2 EmbriologiaDocument6 pagesSemana 3 Parte 2 EmbriologiamarNo ratings yet
- Clase 1 Evolucion y DesarrolloDocument49 pagesClase 1 Evolucion y Desarrolloenfoquepsicosocialnovatos2011100% (1)
- Neuro EmbriologíaDocument17 pagesNeuro EmbriologíaCaleb RodeaNo ratings yet
- Unidad 4 Resumen 1Document8 pagesUnidad 4 Resumen 1abigailNo ratings yet
- 03 Embriologia Tercera A Octava Semana de DesarrolloDocument33 pages03 Embriologia Tercera A Octava Semana de DesarrolloCarlos Kevin Salas DonayreNo ratings yet
- Manual de Neuro Dr. GaribayDocument120 pagesManual de Neuro Dr. GaribayCarla A. CorriloNo ratings yet
- Tercera Semana EmbrionDocument5 pagesTercera Semana EmbrionMar MehhNo ratings yet
- 9-Somitas y NerulaciónDocument3 pages9-Somitas y NerulaciónLuciana RodríguezNo ratings yet
- Clase de Embriología Del SNCDocument19 pagesClase de Embriología Del SNCLuis Angel Unocc GuillenNo ratings yet
- FL-8 de La Tercera A La Octava Semana Del Periodo EmbrionarioDocument12 pagesFL-8 de La Tercera A La Octava Semana Del Periodo EmbrionarioAnonymous tpBeHze3WFNo ratings yet
- Sistema NerviosoDocument18 pagesSistema NerviosoantonioNo ratings yet
- Sistema Nervioso EmbrionarioDocument15 pagesSistema Nervioso EmbrionarioEstefania OchoaNo ratings yet
- Clase 6 Período Embrionario 1Document25 pagesClase 6 Período Embrionario 1Uriel Avendaño100% (1)
- 3º Semana. Gastrulación. Mesodermo y Celoma Intraembrionarios. Neurulación.Document9 pages3º Semana. Gastrulación. Mesodermo y Celoma Intraembrionarios. Neurulación.Notnow MaybelaterNo ratings yet
- Embrión TrilaminarDocument18 pagesEmbrión TrilaminarITZAMARAZAVALANo ratings yet
- Disco Germinativo TrilaminarDocument7 pagesDisco Germinativo TrilaminarJosé Ramón Fernández ChacínNo ratings yet
- Tema 3 EmbriologiaDocument8 pagesTema 3 EmbriologiaJulio SalazarNo ratings yet
- Desarrollo embrionario semana 5Document8 pagesDesarrollo embrionario semana 5Ax3L hushNo ratings yet
- Anatomia y Fisiología del Sistema Nervioso IIFrom EverandAnatomia y Fisiología del Sistema Nervioso IIRating: 4 out of 5 stars4/5 (28)
- 8 1 MamiferosDocument23 pages8 1 MamiferosfernandoNo ratings yet
- 4 2 Caenorhabditis ElegansDocument24 pages4 2 Caenorhabditis ElegansfernandoNo ratings yet
- 8 2 MamiferosDocument12 pages8 2 MamiferosfernandoNo ratings yet
- 8 1 MamiferosDocument23 pages8 1 MamiferosfernandoNo ratings yet
- 4 - 1 - Caenorhabditis Elegans PDFDocument18 pages4 - 1 - Caenorhabditis Elegans PDFfernandoNo ratings yet
- 8 2 MamiferosDocument12 pages8 2 MamiferosfernandoNo ratings yet
- 4 3 Caenorhabditis ElegansDocument19 pages4 3 Caenorhabditis ElegansfernandoNo ratings yet
- 3 Drosophila MelanogasterDocument92 pages3 Drosophila Melanogasterfernando0% (1)
- 4 - 1 - Caenorhabditis Elegans PDFDocument18 pages4 - 1 - Caenorhabditis Elegans PDFfernandoNo ratings yet
- Tercer Parcial 1C 2019Document1 pageTercer Parcial 1C 2019Agus AngeliniNo ratings yet
- Almacenamiento de granos en plantas procesadorasDocument5 pagesAlmacenamiento de granos en plantas procesadorasPëlaïthö Gualli KevinNo ratings yet
- ALEA23CPDocument2 pagesALEA23CPJose Luis Lima BallesterosNo ratings yet
- Evaluación Final 2022-10 FUNDAMENTOS DE PROGRAMACIÓNDocument5 pagesEvaluación Final 2022-10 FUNDAMENTOS DE PROGRAMACIÓNANGELA FERNANDA ROMAN VELIZNo ratings yet
- Introduccion A La Fisica Del Estado Solido CristalografiaDocument75 pagesIntroduccion A La Fisica Del Estado Solido CristalografiaGabriel González GarcíaNo ratings yet
- Tipos de Suelos en El EcuadorDocument29 pagesTipos de Suelos en El EcuadorFreddy XavierNo ratings yet
- PhpMyAdmin SQL DumpDocument7 pagesPhpMyAdmin SQL DumpRobert Jeyson Rivas Sinisterra100% (1)
- Método Vogel flujo aceite gasDocument4 pagesMétodo Vogel flujo aceite gashallibelstark100% (1)
- El Uso de La TecnologíaDocument3 pagesEl Uso de La TecnologíaJuliana LeónNo ratings yet
- Semana 1Document9 pagesSemana 1EduardoNo ratings yet
- Proyecto de Ley #5462/2022-CRDocument12 pagesProyecto de Ley #5462/2022-CRDiario Oficial El PeruanoNo ratings yet
- La Escala Binet-Simon en Estados UnidosDocument1 pageLa Escala Binet-Simon en Estados UnidosHernandez Mendez Monica Lucia 202107160No ratings yet
- Ensayo de PedagogiaDocument11 pagesEnsayo de PedagogiaJuan de la cruz Jimenez HNo ratings yet
- Cimentaciones SuperficialesDocument8 pagesCimentaciones SuperficialesWilsonDELACruzOvandoNo ratings yet
- Sso - Ot.007 Reglamento Interno de SsoDocument48 pagesSso - Ot.007 Reglamento Interno de SsoJordy Barrantes CuadrosNo ratings yet
- Seguridad activa y pasivaDocument24 pagesSeguridad activa y pasivasnake5556100% (1)
- Ventajas y Desventajas de Los PresupuestosDocument26 pagesVentajas y Desventajas de Los PresupuestosJohana OrtízNo ratings yet
- Andres Ysu Nuevo AmigoDocument8 pagesAndres Ysu Nuevo AmigoJuanJoseJimenezAndrade100% (1)
- Unidad de Agosto 2018Document9 pagesUnidad de Agosto 2018Boris WilfredoNo ratings yet
- Procesos psicológicos básicos IIDocument33 pagesProcesos psicológicos básicos IImartinaNo ratings yet
- Costo Obras Drenaje VialDocument9 pagesCosto Obras Drenaje VialGerman Urdaneta H100% (1)
- UntitledDocument53 pagesUntitledSandra CruzNo ratings yet
- Glosario de TerminosDocument5 pagesGlosario de TerminosViticor CarrilloNo ratings yet
- Taller Ofimatica Triptico SaldarriagajoseDocument2 pagesTaller Ofimatica Triptico SaldarriagajoseManuel SaldarriagaNo ratings yet
- Taller Panel de ControlDocument16 pagesTaller Panel de ControlAnonymous 9f9ODWYpNo ratings yet
- PD 11 Sist de Part - Choques 2023-I Fis 1Document4 pagesPD 11 Sist de Part - Choques 2023-I Fis 1DAIVID JHUNIOR ALELUYA UNCHUPAICONo ratings yet
- Fase5 - Trabajo Colaborativo - Grupo - 403010 - 9Document12 pagesFase5 - Trabajo Colaborativo - Grupo - 403010 - 9Mafe LizarazoNo ratings yet
- Informe Del Osciloscopio DigitalDocument9 pagesInforme Del Osciloscopio Digitalfranco_3210_No ratings yet
- 6 1 3 Caso 2 Experiencia 6Document2 pages6 1 3 Caso 2 Experiencia 6Gustavo GarínNo ratings yet
- Ejercicios SecuenciaciónDocument7 pagesEjercicios SecuenciaciónroalNo ratings yet