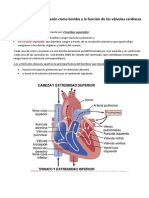
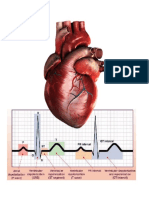
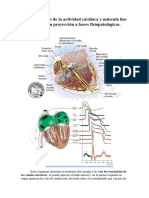
You might also like
- Corazón y Conducción Eléctrica - Parte 1Document3 pagesCorazón y Conducción Eléctrica - Parte 1IgnacioNo ratings yet
- Fisiologia CardiacaDocument17 pagesFisiologia CardiacaGenesis PradaNo ratings yet
- Sistema CardionectorDocument36 pagesSistema CardionectorJosé Carlos Pineda100% (8)
- Fisiologia CardiacaDocument19 pagesFisiologia CardiacaRosa BlancoNo ratings yet
- FCV03. Excitación y Conducción CardíacaDocument37 pagesFCV03. Excitación y Conducción CardíacaSofiaNo ratings yet
- Electrofisiología Del EKGDocument67 pagesElectrofisiología Del EKGHans M. Valdiviezo BonifazNo ratings yet
- Desfibrilador CardiacoDocument46 pagesDesfibrilador CardiacoJesus Resta NavarroNo ratings yet
- MielinizacionDocument57 pagesMielinizacionVictoria Castillo100% (1)
- Músculo CardiacoDocument15 pagesMúsculo CardiacoJose QuinteroNo ratings yet
- Fisiologia CardiacaDocument11 pagesFisiologia CardiacaCarol IsidroNo ratings yet
- Control de Los Patrones de Descarga en El SomaDocument3 pagesControl de Los Patrones de Descarga en El SomaSTEVEN WILTER JUNIOR MENDEZ ALVARADONo ratings yet
- Control de La Función CardiovascularDocument10 pagesControl de La Función CardiovascularMartin Ojeda CarpioNo ratings yet
- Electrofisiologia Cardiaca 1Document9 pagesElectrofisiologia Cardiaca 1Grease Cubas AlzamoraNo ratings yet
- Excitoconductor y EKGDocument16 pagesExcitoconductor y EKGAlbert FigueredoNo ratings yet
- Modelado y Simulación de La Actividad Eléctrica Del CorazonDocument10 pagesModelado y Simulación de La Actividad Eléctrica Del Corazonelalex25No ratings yet
- Sistema CardionectorDocument10 pagesSistema CardionectorNicolas Bravo JimenezNo ratings yet
- Laboratorio 3. Potencial de AccionDocument14 pagesLaboratorio 3. Potencial de AccionMauricio Reyes CastroNo ratings yet
- Biofisica CardíacaDocument28 pagesBiofisica CardíacaMarcos Días100% (1)
- Músculo CardíacoDocument15 pagesMúsculo CardíacoNara FlosanNo ratings yet
- Guía de Estudio 3Document12 pagesGuía de Estudio 3Debora RuizNo ratings yet
- Fisiologia Cardiovascular IDocument92 pagesFisiologia Cardiovascular IMayra Alejandra Gutiérrez SoteloNo ratings yet
- Manual Toma e Interpretacion de ElectrocardiogramaDocument29 pagesManual Toma e Interpretacion de ElectrocardiogramaGuillermina SanchezNo ratings yet
- BiopotencialesDocument24 pagesBiopotencialesMiguel SoberanesNo ratings yet
- Cuestionario de Músculo CardiacoDocument3 pagesCuestionario de Músculo CardiacoJesús Leyva GarcíaNo ratings yet
- Grupo C - Actividad 2cDocument9 pagesGrupo C - Actividad 2cmicacampos.baNo ratings yet
- Mecanismo Del CorazonDocument11 pagesMecanismo Del CorazonJc TovarNo ratings yet
- Ecg CastellanosDocument143 pagesEcg CastellanosVelia Gallegos100% (1)
- Introduccion A La Fisiologia CardiacaDocument16 pagesIntroduccion A La Fisiologia CardiacaMartin PiattiNo ratings yet
- Fisiologia CardiovascularDocument67 pagesFisiologia CardiovascularMirko Huamán CuevaNo ratings yet
- ARRITMIASDocument21 pagesARRITMIASJoselyn RodriguezNo ratings yet
- Cap - 9 (Musculo Cardiaco El Corazon)Document85 pagesCap - 9 (Musculo Cardiaco El Corazon)JONATHAN ARIEL LUNA JIMENEZNo ratings yet
- Corrientes IonicasDocument6 pagesCorrientes Ionicasoscar olivasNo ratings yet
- Resumen Clases EcgDocument8 pagesResumen Clases EcgIsis CruzNo ratings yet
- Bioelectric I DadDocument5 pagesBioelectric I DadAriana Figueroa OviedoNo ratings yet
- Comunicacion InterneuronaDocument13 pagesComunicacion InterneuronaFrank Sanchez MoronNo ratings yet
- Conducción Del Impulso Nervioso Por El AxónDocument16 pagesConducción Del Impulso Nervioso Por El AxónVale LeroyerNo ratings yet
- Practica Potencial de Acción CardiacoDocument9 pagesPractica Potencial de Acción CardiacoXavi MGNo ratings yet
- Parcial 3 (Bioelectricidad)Document16 pagesParcial 3 (Bioelectricidad)Nicole Carolina Quintero RomeroNo ratings yet
- Tarea 2 - Miguel Angel Ahumada Segura!!!Document16 pagesTarea 2 - Miguel Angel Ahumada Segura!!!MIGUEL ANGEL AHUMADANo ratings yet
- Contracción Del Músculo CardíacoDocument11 pagesContracción Del Músculo CardíacoMelanie Acurio OrtizNo ratings yet
- Tipeo Completo Curso ECGDocument51 pagesTipeo Completo Curso ECGVioleta Canales PueblaNo ratings yet
- Tipeo Semana 1 ECGDocument10 pagesTipeo Semana 1 ECGVioleta Canales PueblaNo ratings yet
- Preguntero Con Respuestas - CompletoDocument55 pagesPreguntero Con Respuestas - CompletoFranciscoMolinaNo ratings yet
- Canales Iónicos - Qué Son, Tipos. y Cómo Funcionan en Las CélulasDocument19 pagesCanales Iónicos - Qué Son, Tipos. y Cómo Funcionan en Las CélulasBruceNo ratings yet
- ELECTROCARDIOGRAFIADocument113 pagesELECTROCARDIOGRAFIArosaNo ratings yet
- Electrofisiologia Cardiaca2022Document53 pagesElectrofisiologia Cardiaca2022yoyoyoyoNo ratings yet
- FisiologiaDocument11 pagesFisiologiaMarjorie PérezNo ratings yet
- MECDocument2 pagesMECGerhard VazquezNo ratings yet
- Cuestionario Biología 'Impulso Nervioso'Document1 pageCuestionario Biología 'Impulso Nervioso'Tomás Pérez AmentaNo ratings yet
- Fisiologia de La SinapsisDocument33 pagesFisiologia de La SinapsisThalia Aguilar OrtegaNo ratings yet
- Fisiología - Guía # 1. Actividad Electrica Cardiaca 2023-2Document10 pagesFisiología - Guía # 1. Actividad Electrica Cardiaca 2023-2LAURA SOFIA PEREZ CUELLONo ratings yet
- Documento Sin TítuloDocument15 pagesDocumento Sin TítuloGuillermina SanchezNo ratings yet
- Fisiología Cardiovascular Electrocardiografía N°4 PDFDocument22 pagesFisiología Cardiovascular Electrocardiografía N°4 PDFKevin Argotti CazarNo ratings yet
- Potencial de Accion CardiacoDocument8 pagesPotencial de Accion CardiacoAngelo Anaya CaponeNo ratings yet
- CARDIO - Bases Biofísicas de La Actividad Cardiaca y Músculo Liso Vascular Con Proyección A Bases FisiopatológicasDocument28 pagesCARDIO - Bases Biofísicas de La Actividad Cardiaca y Músculo Liso Vascular Con Proyección A Bases FisiopatológicasRomina NogueiraNo ratings yet
- Cuestionario Fisio IIDocument9 pagesCuestionario Fisio IIEsteban RobloxNo ratings yet
- Experiencias sobre circuitos eléctricos en serie operados por corriente directaFrom EverandExperiencias sobre circuitos eléctricos en serie operados por corriente directaNo ratings yet
- EstudiosDocument2 pagesEstudiosSimón WitonNo ratings yet
- Ley Nacional de FolkloreDocument4 pagesLey Nacional de FolkloreSimón WitonNo ratings yet
- Balance Glomerulotubular Retroalimentacion TubuloglomerularDocument22 pagesBalance Glomerulotubular Retroalimentacion TubuloglomerularSimón WitonNo ratings yet
- Atlas de Anatomía Depto Anatomía FMed UBADocument202 pagesAtlas de Anatomía Depto Anatomía FMed UBALaura Panique LazcanoNo ratings yet
- Mdicamentos CitostticosDocument15 pagesMdicamentos CitostticosSimón WitonNo ratings yet
- Casos Clinicos 2012 13 29junio PDFDocument85 pagesCasos Clinicos 2012 13 29junio PDFSimón WitonNo ratings yet
- 2 Relación Médico-PacienteDocument16 pages2 Relación Médico-PacienteSimón WitonNo ratings yet
- Recuperatorio 2 Parcial BQ Fmed UbaDocument4 pagesRecuperatorio 2 Parcial BQ Fmed UbaSimón WitonNo ratings yet
- Proyecto Rio Blanco Problemas AmbientalesDocument15 pagesProyecto Rio Blanco Problemas AmbientalesJose PantaNo ratings yet
- Suscripción Digital - Todas Las Noticias de Colombia en Un Lugar - EL ESPECTADORDocument2 pagesSuscripción Digital - Todas Las Noticias de Colombia en Un Lugar - EL ESPECTADORalfonsoNo ratings yet
- Resolucion 2020060113363 Zonas Dificil Acceso 2021Document78 pagesResolucion 2020060113363 Zonas Dificil Acceso 2021hugoivNo ratings yet
- Valoración de Dos Sistemas Acuapónicos para El Cultivo de FresaDocument78 pagesValoración de Dos Sistemas Acuapónicos para El Cultivo de FresaBramyer Rafael Arias PortilloNo ratings yet
- Exposicion EncuestaDocument8 pagesExposicion EncuestaDiaz Pacheco OyukiNo ratings yet
- Sopa de Letras de Ciencias Auxiliares de La HistoriaDocument1 pageSopa de Letras de Ciencias Auxiliares de La HistoriaUlises TipaNo ratings yet
- Constancia de Conformidad de Servicio 2Document2 pagesConstancia de Conformidad de Servicio 2Oskar VelaNo ratings yet
- TesisDocument157 pagesTesisEvangelina GonzalezNo ratings yet
- Diagrama BimanualDocument2 pagesDiagrama BimanualWaldo Colón BejaránNo ratings yet
- Procedimiento de Riesgo PsicosocialDocument4 pagesProcedimiento de Riesgo PsicosocialS'ebas P'sNo ratings yet
- Diapositivas Monarquia y AbsolutismoDocument39 pagesDiapositivas Monarquia y AbsolutismoCarlos Enrique Barzola PalominoNo ratings yet
- Tarea 8 Introduccion A La AgrimensuraDocument6 pagesTarea 8 Introduccion A La AgrimensuraIsaac TaverasNo ratings yet
- Papel Tapiz-Guia de Tipos y UsosDocument17 pagesPapel Tapiz-Guia de Tipos y UsosEdgar MirandaNo ratings yet
- 2.1 - Adhesio - N de Las Resinas Compuestas A Los Tejidos Dentarios BertoldiDocument46 pages2.1 - Adhesio - N de Las Resinas Compuestas A Los Tejidos Dentarios BertoldiMaximiliano HidalgoNo ratings yet
- Guia Helipuertos PDFDocument9 pagesGuia Helipuertos PDFSeiya Neyogiri NgupiNo ratings yet
- Desarrollo Lógico de La InformaciónDocument2 pagesDesarrollo Lógico de La InformaciónNoraPintosPerrottaNo ratings yet
- Ampliacion LenguaDocument24 pagesAmpliacion LenguaGuacimara Garcia GutierrezNo ratings yet
- Clase10 - Soluciones RealesDocument30 pagesClase10 - Soluciones RealesSofíaGuajardoNo ratings yet
- 33 - Edición-Técnica-Pintura-de-Alto-Tráfico-Recomendaciones-para-la-Selección-y-Aplicación-en-Recubrimientos-de-PisoDocument43 pages33 - Edición-Técnica-Pintura-de-Alto-Tráfico-Recomendaciones-para-la-Selección-y-Aplicación-en-Recubrimientos-de-Pisochinese0608No ratings yet
- Diferencia Entre Croquis y MapaDocument1 pageDiferencia Entre Croquis y Mapaoscardwpearl33% (6)
- Probabilidad y EstadisticaDocument4 pagesProbabilidad y EstadisticaLisi ReyesNo ratings yet
- Marleny 1Document14 pagesMarleny 1Cristian JoseNo ratings yet
- Experiencia 01-145Document21 pagesExperiencia 01-145Flor Vega VasquezNo ratings yet
- Article Como Hackear Un FacebookDocument2 pagesArticle Como Hackear Un Facebookmaggie2orr60No ratings yet
- Valores Normales de LaboratorioDocument4 pagesValores Normales de LaboratorioMelanieNo ratings yet
- Pre InformeDocument24 pagesPre InformeJoceline Murguia ChamasNo ratings yet
- Jesús Ante Pilato Desde La Matríz Social TriádicaDocument12 pagesJesús Ante Pilato Desde La Matríz Social TriádicaMiguel La RottaNo ratings yet
- PUTUMAYO ENSAYO FNDocument4 pagesPUTUMAYO ENSAYO FNAlfonsina Quiroga VargasNo ratings yet
- Como Curar La Eyaculación PrecozDocument9 pagesComo Curar La Eyaculación PrecozAlex Hernández ToroNo ratings yet
- Pasos Implementaciòn de Un Sistema de Gestion de La CalidadDocument5 pagesPasos Implementaciòn de Un Sistema de Gestion de La CalidadBRAHIAN STEVEN PARRADO CADENANo ratings yet