You might also like
- ZOOLOGÍA DE VERTEBRADOS Clase 4 Peces IIDocument47 pagesZOOLOGÍA DE VERTEBRADOS Clase 4 Peces IIJordan RuizNo ratings yet
- Bioquimica Devlin 4a EdicionDocument621 pagesBioquimica Devlin 4a EdicionLuisNo ratings yet
- Temas Examen de Bioquímica IIDocument9 pagesTemas Examen de Bioquímica IIManuel Alejandro Iza GutierrezNo ratings yet
- Histologia Basica Texto y Atlas Junqueira CarneiroDocument557 pagesHistologia Basica Texto y Atlas Junqueira CarneiroDanna Alducin0% (1)
- Aplicaciones de La Formula de HendersonDocument3 pagesAplicaciones de La Formula de HendersonClaudymar GonzalezNo ratings yet
- 4) Rivolta, M. y Benavides, L. (2017), Apunte de Cátedra Unidad 4. Bases Fisicoquimicas de La Vida PDFDocument12 pages4) Rivolta, M. y Benavides, L. (2017), Apunte de Cátedra Unidad 4. Bases Fisicoquimicas de La Vida PDFMauricio Exequiel Cazado TorassoNo ratings yet
- Señalizacion Celular PDFDocument48 pagesSeñalizacion Celular PDFOscar Ivan0% (1)
- Fisiopatología - Porth - 7ed PDFDocument1,568 pagesFisiopatología - Porth - 7ed PDFJr Adrian Bedregal Gomez90% (10)
- Apuntes Biologia Celular y TisularDocument152 pagesApuntes Biologia Celular y TisularFatima Jerez50% (2)
- Microbiologia Médica - BasualdoDocument1,425 pagesMicrobiologia Médica - BasualdoEnrique Duarte81% (21)
- Bioquimica de Ferrier PDFDocument531 pagesBioquimica de Ferrier PDFCarolina Gómez Mora81% (16)
- Lehninger Principios de Bioquimica 7a EdicionDocument1,235 pagesLehninger Principios de Bioquimica 7a EdicionJuan Pablo Suarez Morales100% (1)
- Biologia e Introduccion A La Biologia CelularDocument128 pagesBiologia e Introduccion A La Biologia CelularAngie EstradaNo ratings yet
- Bioquímica 200-2645Document15 pagesBioquímica 200-2645bioquimica100% (1)
- LehningerDocument1,283 pagesLehningerMelissa DiazNo ratings yet
- Trabajo HemoglobinaDocument3 pagesTrabajo HemoglobinaAlejandroToqueNo ratings yet
- Amortiguadores SanguíneosDocument3 pagesAmortiguadores SanguíneosAna Alejandra Deluque CampoNo ratings yet
- CURTIS Uba Xxi 2021 7ma EdDocument135 pagesCURTIS Uba Xxi 2021 7ma Edjessica carolineNo ratings yet
- Classificacio EnzimsDocument3 pagesClassificacio EnzimsjbuedosNo ratings yet
- Metabolismo: conceptos básicos y visión de conjuntoDocument23 pagesMetabolismo: conceptos básicos y visión de conjuntoAlexis Naula100% (1)
- Cuando Se Analiza Un DNA PlasmídicoDocument1 pageCuando Se Analiza Un DNA PlasmídicoLeydi Gutierrez ArmijosNo ratings yet
- Sinergia y RecursividadDocument35 pagesSinergia y RecursividadNit_suNo ratings yet
- Acidos y BasesDocument9 pagesAcidos y BasesKATHERYN SUESCUN GUEVARA100% (1)
- Principios de Bioquímica Lehninger - David L. Nelson, Michael M. Cox - 5ta Edición PDFDocument1,283 pagesPrincipios de Bioquímica Lehninger - David L. Nelson, Michael M. Cox - 5ta Edición PDFThematic BoxNo ratings yet
- Bioquímica (Stryer) 6ta EdiciónDocument1,137 pagesBioquímica (Stryer) 6ta EdiciónFederico Ariganello17% (6)
- El Hígado (Semiologia)Document24 pagesEl Hígado (Semiologia)Enrique Ollarbe Castillo100% (1)
- Manual de Fisiologia (UNAM)Document115 pagesManual de Fisiologia (UNAM)Alfonso AcevedoNo ratings yet
- Uniones IntercelularesDocument27 pagesUniones IntercelularesSusana Salas NuñezNo ratings yet
- Bioquímica Médica (4a. Ed.) - (18 Metabolismo de Las Lipoproteínas y Aterogénesis)Document22 pagesBioquímica Médica (4a. Ed.) - (18 Metabolismo de Las Lipoproteínas y Aterogénesis)Ximena Miranda SalgadoNo ratings yet
- Peptidos de Importancia BiologicaDocument12 pagesPeptidos de Importancia BiologicaPaola MoraNo ratings yet
- Pierce Un Enfoque Conceptual GeneticaDocument617 pagesPierce Un Enfoque Conceptual GeneticaCarlos Zepeda Guido50% (4)
- Comunicación celular indirectaDocument3 pagesComunicación celular indirectaDaniel Romano100% (4)
- Investigues de Micro Arreglado 1Document6 pagesInvestigues de Micro Arreglado 1Albert Diaz100% (8)
- Origen y Evolución de La Celula PDFDocument40 pagesOrigen y Evolución de La Celula PDFEmmanuel Jaramillo Orozco50% (2)
- Bioquímica: Conceptos Esenciales. 3 EdiciónDocument2 pagesBioquímica: Conceptos Esenciales. 3 Ediciónsharon perez0% (1)
- Monografia Del Ovulo, JhackDocument2 pagesMonografia Del Ovulo, JhackJhackBaltazarMendozaNo ratings yet
- ORD SUPLENTE Examen LenguaCastellana LiteraturaIIDocument3 pagesORD SUPLENTE Examen LenguaCastellana LiteraturaIIRuidong LinNo ratings yet
- 1 Hipotalamo Hipofisis-Completa 2017Document49 pages1 Hipotalamo Hipofisis-Completa 2017virginiaNo ratings yet
- Resumen Ross Paco Reynoso Haro HECHO UDG...Document54 pagesResumen Ross Paco Reynoso Haro HECHO UDG...Cecilia Michelle RomoNo ratings yet
- FisiologiaDocument17 pagesFisiologiaCristian CastellonNo ratings yet
- Panamericana Anatomia Humana en Casos Clinicos 5a Edicion 856124Document4 pagesPanamericana Anatomia Humana en Casos Clinicos 5a Edicion 856124Jeshue BarrazaNo ratings yet
- 44 BioquimicaDocument206 pages44 BioquimicaIrma Ruiz94% (31)
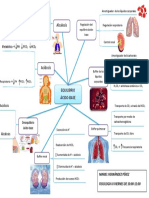
- Regulación del equilibrio ácido-base a través de los sistemas respiratorio y renalDocument1 pageRegulación del equilibrio ácido-base a través de los sistemas respiratorio y renalMariel Hernândez Pêrez100% (1)
- Tipos y efecto de mutacionesDocument9 pagesTipos y efecto de mutacionesAlejandro SilvaNo ratings yet
- Bohinski BioquimicaDocument763 pagesBohinski BioquimicaAlejandra Baas Guzmán100% (3)
- Difusion y OsmosisDocument8 pagesDifusion y OsmosisAndres BolivarNo ratings yet
- BERNE y LEVY. FisiologíaDocument11 pagesBERNE y LEVY. FisiologíaCarolina Barrio0% (1)
- Apreciación Del Poder Regulador de Las Soluciones Buffer y El Efecto de La ConcentraciónDocument8 pagesApreciación Del Poder Regulador de Las Soluciones Buffer y El Efecto de La ConcentraciónXochitlNo ratings yet
- Estructura y circunvoluciones del telencéfaloDocument5 pagesEstructura y circunvoluciones del telencéfaloangeldeluz_morena1390No ratings yet
- Hematoxilina PDFDocument51 pagesHematoxilina PDFthalia0% (1)
- Recursos para la enseñanza/aprendizaje del metabolismoFrom EverandRecursos para la enseñanza/aprendizaje del metabolismoNo ratings yet
- Bioquimica Tercera Edicion Libro de TextDocument6 pagesBioquimica Tercera Edicion Libro de TextAracely PeñaNo ratings yet
- Traducción: Transcripción, Traduc Ción Y Replicación CelularDocument1 pageTraducción: Transcripción, Traduc Ción Y Replicación CelularNtuys ChNo ratings yet
- Victoria Sanchez 9C Estructura Del AdnDocument8 pagesVictoria Sanchez 9C Estructura Del AdnVictoria Sanchez NietoNo ratings yet
- Organizador Grafico Información GenéticaDocument3 pagesOrganizador Grafico Información Genéticalquinonez9760No ratings yet
- Código Genético y Síntesis de ProteínasDocument15 pagesCódigo Genético y Síntesis de ProteínasMariana100% (1)
- Procesos de Iniciación de Cadena Rezagada en La Replicación de ADN de EucariotasDocument3 pagesProcesos de Iniciación de Cadena Rezagada en La Replicación de ADN de EucariotasAdriana Vanesa Tueros GuzmánNo ratings yet
- Bipiav9g 10Document9 pagesBipiav9g 10Milton Edward Robles ArquerosNo ratings yet
- Aplicación de Bases Moelculares PDFDocument2 pagesAplicación de Bases Moelculares PDFPao SotoNo ratings yet
- Presentación AcidosNucleicosDocument17 pagesPresentación AcidosNucleicosEdith DanielNo ratings yet
- RVM #235-2021-MineduDocument220 pagesRVM #235-2021-MineduvilmaNo ratings yet
- 2021 Motivacion LaboralDocument20 pages2021 Motivacion LaboralwaldirNo ratings yet
- Trabajos en Altura 07-08Document32 pagesTrabajos en Altura 07-08waldirNo ratings yet
- Nectar de LimonDocument35 pagesNectar de LimonwaldirNo ratings yet
- Yo Soy Abundancia Patricia AnayaDocument220 pagesYo Soy Abundancia Patricia AnayaJOctavioMuñozReyesNo ratings yet
- Yo Soy Abundancia Patricia AnayaDocument220 pagesYo Soy Abundancia Patricia AnayaJOctavioMuñozReyesNo ratings yet
- Modelo de negocio CanvasDocument112 pagesModelo de negocio CanvasOscar Roberto Recinos Cañas100% (9)
- PSU Matemática: 530 preguntas de facsímiles oficialesDocument85 pagesPSU Matemática: 530 preguntas de facsímiles oficialesJessica Riquelme100% (2)
- Encuesta de Residuos SolidosDocument3 pagesEncuesta de Residuos SolidoswaldirNo ratings yet
- Proyecto de Mantenimiento de Vivero MunicipalDocument35 pagesProyecto de Mantenimiento de Vivero MunicipalAyelinNo ratings yet
- Norma Codex para zumos y néctares de frutasDocument21 pagesNorma Codex para zumos y néctares de frutasLuis Torres100% (1)
- Guia Tecnica de Viveros y Semilleros FrutalesDocument40 pagesGuia Tecnica de Viveros y Semilleros FrutalesFabian Cruz R100% (1)
- Produ To Send o XenosDocument100 pagesProdu To Send o XenosJavi Quispillo HuamánNo ratings yet
- Pauta - Planes - de - Negocio - Cafe - PROCOMPITE PDFDocument140 pagesPauta - Planes - de - Negocio - Cafe - PROCOMPITE PDFDenisse Trudy Ingaroca FabianNo ratings yet
- Como Comenzar Un Negocio de Exportación PROMPERUDocument35 pagesComo Comenzar Un Negocio de Exportación PROMPERUGabriela K. Vilca CruzNo ratings yet
- Modulo 1 - Tema 1 - Introducción A LS - DYNADocument8 pagesModulo 1 - Tema 1 - Introducción A LS - DYNAAnonymous siROwVWXNo ratings yet
- Argentina: Derechos arancelarios y otros impuestos a las importacionesDocument6 pagesArgentina: Derechos arancelarios y otros impuestos a las importacioneswaldirNo ratings yet
- Operaciones BancariasDocument26 pagesOperaciones BancariaswaldirNo ratings yet
- Modulo 1 - Tema 2. Tipos de Modelización Con LS-DYNA I PDFDocument6 pagesModulo 1 - Tema 2. Tipos de Modelización Con LS-DYNA I PDFwaldirNo ratings yet
- Modulo 1 - Tema 2. Tipos de Modelización Con LS-DYNA I PDFDocument6 pagesModulo 1 - Tema 2. Tipos de Modelización Con LS-DYNA I PDFwaldirNo ratings yet
- NutricionDocument14 pagesNutricionwayaveraNo ratings yet
- DESHIDRATACIONDocument140 pagesDESHIDRATACIONLanders Salas RolandoNo ratings yet
- Modelos de OligopolioDocument10 pagesModelos de OligopolioCristian MoratayaNo ratings yet
- Modulo 1 - Tema 2. Tipos de Modelización Con LS-DYNA IDocument12 pagesModulo 1 - Tema 2. Tipos de Modelización Con LS-DYNA IwaldirNo ratings yet
- Separata Pulpas Nèctares, Merm Desh, Osmodes y Fruta ConfitadaDocument61 pagesSeparata Pulpas Nèctares, Merm Desh, Osmodes y Fruta ConfitadaLuis MidWan100% (1)
- Colombia Aranceles y Politica ComercialDocument3 pagesColombia Aranceles y Politica ComercialwaldirNo ratings yet
- FAQ Mód1 EstadisticaDocument9 pagesFAQ Mód1 EstadisticaJorshinio Salazar CastilloNo ratings yet
- Importaciones en Perú: marco legal y tipos según transacciónDocument54 pagesImportaciones en Perú: marco legal y tipos según transacciónwaldirNo ratings yet
- Técnicas de Procesado de CarnesDocument17 pagesTécnicas de Procesado de CarnesDarwinVargasMantillaNo ratings yet
- La influencia de la velocidad de agitación en la lixiviación dinámicaDocument8 pagesLa influencia de la velocidad de agitación en la lixiviación dinámicaNano NandoNo ratings yet
- Cuestionarios Laboratorio de BiologíaDocument9 pagesCuestionarios Laboratorio de BiologíaSuellen Gi Mo PiNo ratings yet
- Macronutrientes y MicronutrientesDocument2 pagesMacronutrientes y MicronutrientesArianne Villicaña0% (1)
- Mat4-U2-Sesion 09 2017Document7 pagesMat4-U2-Sesion 09 2017Oscar Jesús Piñas VivasNo ratings yet
- Planta de Producción de OvoproductosDocument162 pagesPlanta de Producción de OvoproductosedgarNo ratings yet
- Metabolismo de ProteinasDocument61 pagesMetabolismo de ProteinasprietalindaNo ratings yet
- Moleculas de La VidaDocument13 pagesMoleculas de La VidaJose vladimir Vela perézNo ratings yet
- Caseinato de SodioDocument9 pagesCaseinato de SodioMarlene JuárezNo ratings yet
- Trabajo 1 Analisis de AlimentosDocument8 pagesTrabajo 1 Analisis de AlimentosAngélica JiménezNo ratings yet
- Gregor MendelDocument3 pagesGregor Mendelcesar vasquezNo ratings yet
- BromatologiaDocument6 pagesBromatologiasergiof vegaNo ratings yet
- Tema 1 (Tamaño, Forma, Volumen)Document24 pagesTema 1 (Tamaño, Forma, Volumen)Mario Enríquez0% (1)
- Qui U2 Ea CLRGDocument6 pagesQui U2 Ea CLRGClaudia RomeroNo ratings yet
- Mapa ConceptualDocument3 pagesMapa ConceptualJairo TavaresNo ratings yet
- 6 Determinacion FactorDocument3 pages6 Determinacion FactorYoselin GomezNo ratings yet
- 10 1007@s00438-002-0702-5 en EsDocument11 pages10 1007@s00438-002-0702-5 en EsKarla Edith CortesNo ratings yet
- Clasificación de Las ProteínasDocument3 pagesClasificación de Las ProteínasZulyNo ratings yet
- Biología 1 - Examen PDFDocument2 pagesBiología 1 - Examen PDFLorena Soriano HernandezNo ratings yet
- Manual de LaboratorioDocument45 pagesManual de LaboratorioPabloNo ratings yet
- Valor Nutritivo de Los AlimentosDocument8 pagesValor Nutritivo de Los AlimentosMaximiliano Solis100% (1)
- Muerte Microbiana Por Agentes Fisicos y QuimicosDocument8 pagesMuerte Microbiana Por Agentes Fisicos y QuimicosIvanna Andrea CharrisNo ratings yet
- HistidinaDocument10 pagesHistidinaDayana DumancelaNo ratings yet
- Tesis Suero de LecheDocument155 pagesTesis Suero de LecheVictor Burgos50% (2)
- PRACTICA 3-Cuantifica ProteinasDocument10 pagesPRACTICA 3-Cuantifica ProteinasAlvaro JesusNo ratings yet
- Cartel de ProteinasDocument1 pageCartel de ProteinasROSA100% (1)
- Apunte 4Document10 pagesApunte 4Lara ReevesNo ratings yet
- Propiedades Quimicas y Determinación Del Pi de Las ProteinasDocument17 pagesPropiedades Quimicas y Determinación Del Pi de Las ProteinascarlosNo ratings yet
- Química cosméticos cabelloDocument7 pagesQuímica cosméticos cabelloClaudiaRosaPonceChapoñan100% (2)
- Mod. de Nutrientes.2022Document66 pagesMod. de Nutrientes.2022ARIANA LISBETH ZAVALA MIRANDANo ratings yet
- Cuadernillo 2Document31 pagesCuadernillo 2Fredy Alba100% (1)
- Produccion de CodornicesDocument26 pagesProduccion de CodorniceszairaNo ratings yet