You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Method Statement For Lifting WorksDocument12 pagesMethod Statement For Lifting WorksRachel Flores85% (26)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Fortune Falls - Lou VaneDocument402 pagesFortune Falls - Lou VanetesteNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Estrogen Dominance-The Silent Epidemic by DR Michael LamDocument39 pagesEstrogen Dominance-The Silent Epidemic by DR Michael Lamsmtdrkd75% (4)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Full Carrino Plaza Brochure and Application (General)Document8 pagesFull Carrino Plaza Brochure and Application (General)tanis581No ratings yet
- Margarita ForesDocument20 pagesMargarita ForesKlarisse YoungNo ratings yet
- MGMTDocument19 pagesMGMTMakrant MohanNo ratings yet
- NT PotentialMiningDevelopmentsDocument13 pagesNT PotentialMiningDevelopmentsho100hoNo ratings yet
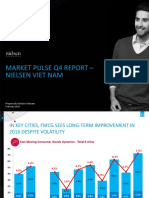
- Market Pulse Q4 Report - Nielsen Viet Nam: Prepared by Nielsen Vietnam February 2017Document8 pagesMarket Pulse Q4 Report - Nielsen Viet Nam: Prepared by Nielsen Vietnam February 2017K57.CTTT BUI NGUYEN HUONG LYNo ratings yet
- W4. Grade 10 Health - Q1 - M4 - v2Document22 pagesW4. Grade 10 Health - Q1 - M4 - v2Jesmael PantalunanNo ratings yet
- Withania DSC PDFDocument6 pagesWithania DSC PDFEfsha KhanNo ratings yet
- Jinko 570 Mono Facial Jkm570m-7rl4-VDocument2 pagesJinko 570 Mono Facial Jkm570m-7rl4-VShahneela AnsariNo ratings yet
- 365 Days (Blanka Lipińska)Document218 pages365 Days (Blanka Lipińska)rjalkiewiczNo ratings yet
- Intimacy and Healthy Affective Maturaity - Fa-Winter09bDocument9 pagesIntimacy and Healthy Affective Maturaity - Fa-Winter09bCarlos GiraldoNo ratings yet
- MSDS of Poly Aluminum ChlorideDocument5 pagesMSDS of Poly Aluminum ChlorideGautamNo ratings yet
- 351 UN 1824 Sodium Hydroxide SolutionDocument8 pages351 UN 1824 Sodium Hydroxide SolutionCharls DeimoyNo ratings yet
- Impact of Job Design On Employee Engagement: A Theoretical and Literature ReviewDocument6 pagesImpact of Job Design On Employee Engagement: A Theoretical and Literature ReviewAnonymous CwJeBCAXpNo ratings yet
- Uni of Glasgow Map PDFDocument1 pageUni of Glasgow Map PDFJiaying SimNo ratings yet
- Plastic As Soil StabilizerDocument28 pagesPlastic As Soil StabilizerKhald Adel KhaldNo ratings yet
- Teleperformance Global Services Private Limited: Full and Final Settlement - December 2023Document3 pagesTeleperformance Global Services Private Limited: Full and Final Settlement - December 2023vishal.upadhyay9279No ratings yet
- Laboratory Medicine Internship BookletDocument95 pagesLaboratory Medicine Internship BookletMuhammad Attique100% (1)
- An Enhanced Model of Thermo Mechanical Loading On A Vaccum Insulated GlazingDocument29 pagesAn Enhanced Model of Thermo Mechanical Loading On A Vaccum Insulated GlazingNguyễn SơnNo ratings yet
- The Coca-Cola Company - Wikipedia, The Free EncyclopediaDocument11 pagesThe Coca-Cola Company - Wikipedia, The Free EncyclopediaAbhishek ThakurNo ratings yet
- We Find The Way: Shipping InstructionsDocument10 pagesWe Find The Way: Shipping InstructionsLuke WangNo ratings yet
- DAMPNESSDocument21 pagesDAMPNESSChukwu SolomonNo ratings yet
- 1Manuscript-BSN-3y2-1A-CEDILLO-222 11111Document32 pages1Manuscript-BSN-3y2-1A-CEDILLO-222 11111SHARMAINE ANNE POLICIOSNo ratings yet
- WCC PSV PDFDocument40 pagesWCC PSV PDFAlejandro RamelaNo ratings yet
- Phoenix Contact DATA SHEETDocument16 pagesPhoenix Contact DATA SHEETShivaniNo ratings yet
- Factors That Contribute To Successful BakingDocument8 pagesFactors That Contribute To Successful BakingErrol San Juan100% (1)
- R. Nishanth K. VigneswaranDocument20 pagesR. Nishanth K. VigneswaranAbishaTeslinNo ratings yet
- Plastic Omnium 2015 RegistrationDocument208 pagesPlastic Omnium 2015 Registrationgsravan_23No ratings yet