You might also like
- Manual Bioquímica Clínica UNAMDocument160 pagesManual Bioquímica Clínica UNAMNeo Aqp100% (1)
- Caracteristicas de Desinfectantes Seleccionados 2016Document1 pageCaracteristicas de Desinfectantes Seleccionados 2016Nazaret BaskervilleNo ratings yet
- Examen San Marcos 2015Document61 pagesExamen San Marcos 2015Zona PreNo ratings yet
- Secuencia Didáctica - La CélulaDocument15 pagesSecuencia Didáctica - La CélulaTasha Duncan100% (1)
- El Derecho GenéticoDocument2 pagesEl Derecho GenéticoGza ChrisNo ratings yet
- EspecieDocument51 pagesEspecieJoseNo ratings yet
- Unidad 12Document3 pagesUnidad 12alvaroNo ratings yet
- Biografia PasteurDocument3 pagesBiografia PasteurJoseNo ratings yet
- Radicales LibresDocument4 pagesRadicales LibresJoseNo ratings yet
- Gases IdealesDocument9 pagesGases IdealesJoseNo ratings yet
- 4 Caida Libre y Tiro Vertical 9 Páginas PDFDocument9 pages4 Caida Libre y Tiro Vertical 9 Páginas PDFJamesgros28No ratings yet
- Energia Interna Gases Ideales PDFDocument14 pagesEnergia Interna Gases Ideales PDFandreinalicNo ratings yet
- Gases IdealesDocument12 pagesGases IdealesPatricia Chambergo RuizNo ratings yet
- Caida LibreDocument7 pagesCaida LibreElgran PoderosoNo ratings yet
- 1606091437.DICOTILEDONEAS Parte - I PDFDocument29 pages1606091437.DICOTILEDONEAS Parte - I PDFleidy dani parraNo ratings yet
- Simple FutureDocument5 pagesSimple FutureJoseNo ratings yet
- Uniones IntercelularesDocument25 pagesUniones IntercelularesJoseNo ratings yet
- Movimiento de Caida Libre 0 PDFDocument8 pagesMovimiento de Caida Libre 0 PDFJagagaaNo ratings yet
- MruDocument6 pagesMruEdwin Chan ChanNo ratings yet
- Movimiento parabólico de caída libre (MPCLDocument2 pagesMovimiento parabólico de caída libre (MPCLJoseNo ratings yet
- Ciclo Celular MitosisDocument0 pagesCiclo Celular MitosisGuadalupeNo ratings yet
- Mec Lab06Document4 pagesMec Lab06Anonymous Dje2UbsjNo ratings yet
- Canales iónicos y potencial de membranaDocument6 pagesCanales iónicos y potencial de membranaJoseNo ratings yet
- Reglamento2018-II NX PDFDocument78 pagesReglamento2018-II NX PDFDaniel Ivan Jo PunNo ratings yet
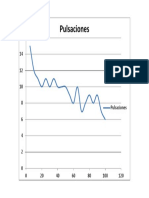
- Gráfica de PulsacionesDocument1 pageGráfica de PulsacionesJoseNo ratings yet
- Problemas de Porcentajes 2 - Eso PDFDocument4 pagesProblemas de Porcentajes 2 - Eso PDFAlan CovarrubiasNo ratings yet
- Leyes de MendelDocument9 pagesLeyes de MendelOrlandd SanchezzNo ratings yet
- Potencial de Acción NerviosoDocument21 pagesPotencial de Acción NerviosoJoseNo ratings yet
- TaxonomyDocument64 pagesTaxonomyJoseNo ratings yet
- Capítulo 3 Mantenimiento Del Volumen CelularDocument9 pagesCapítulo 3 Mantenimiento Del Volumen CelularJoseNo ratings yet
- Capítulo 5 Potencial de MembranaDocument15 pagesCapítulo 5 Potencial de MembranaJoseNo ratings yet
- 4-Ligamiento y RecombinacionDocument17 pages4-Ligamiento y RecombinacionJefferson Moreno100% (1)
- Guia Completa 260-2004Document77 pagesGuia Completa 260-2004Marcelo TorresNo ratings yet
- Microbiologia PDFDocument27 pagesMicrobiologia PDFKeyko Sofia De La Cruz PizanNo ratings yet
- Folleto Celula FinalDocument2 pagesFolleto Celula Finalcarlos javier parra rubio100% (2)
- Introducción de La MitosisDocument2 pagesIntroducción de La MitosisJuan Espinoza CcoraNo ratings yet
- Boramin CA Ficha TecnicaDocument1 pageBoramin CA Ficha TecnicaAlejandro Osorio ToroNo ratings yet
- Fermentacion 6 (Retencion de Gas y Acido en Las Masas)Document2 pagesFermentacion 6 (Retencion de Gas y Acido en Las Masas)Luis MartinNo ratings yet
- El Ciclo Celular - Mitosis y MeiosisDocument9 pagesEl Ciclo Celular - Mitosis y MeiosisJose AristiguietaNo ratings yet
- Cuestionariode BM 5-6Document6 pagesCuestionariode BM 5-6Betsi Emperatriz Arroyo VilaNo ratings yet
- Caracteristicas de Los Microorganismos IndustrialesDocument5 pagesCaracteristicas de Los Microorganismos IndustrialesStephanieMontalvoNo ratings yet
- Los Nucleótidos Son Los Bloques de Construcción de Los Ácidos NucleicosDocument3 pagesLos Nucleótidos Son Los Bloques de Construcción de Los Ácidos NucleicosJennifer RosaNo ratings yet
- Definiciones para El Tema de Las Leyes de MendelDocument3 pagesDefiniciones para El Tema de Las Leyes de MendelRamon Aragon100% (1)
- Funciones del núcleo celularDocument14 pagesFunciones del núcleo celularmprm2579No ratings yet
- 8 - Marcadores GenéticosDocument42 pages8 - Marcadores GenéticoschristianferNo ratings yet
- NeurogenesisDocument5 pagesNeurogenesisbarbaraNo ratings yet
- Aislamiento de bacteriófagos de fuentes naturalesDocument6 pagesAislamiento de bacteriófagos de fuentes naturalesAngel BeltranNo ratings yet
- Medios de Cultivos Mohos y LevadurasDocument2 pagesMedios de Cultivos Mohos y LevadurasLuzNo ratings yet
- 32 Ferm Acetica TitulacionDocument3 pages32 Ferm Acetica TitulacionAngélica R.No ratings yet
- Tecnología de ADN RecombinanteDocument24 pagesTecnología de ADN Recombinantefsilvaf93No ratings yet
- Desarrollo humano y morfología molecularDocument2 pagesDesarrollo humano y morfología molecularCarolina UrreaNo ratings yet
- Plan BQMDocument1 pagePlan BQMNamie MirandaNo ratings yet
- Informe de Morfologia BacterianaDocument4 pagesInforme de Morfologia BacterianaAstrid Êspalza50% (2)
- 6 Taller de Acidos NucleicosDocument5 pages6 Taller de Acidos NucleicosRôdríìgô Dmz Exziite50% (4)
- Reproduccion y Muerte CelularDocument18 pagesReproduccion y Muerte CelularRobert Carvajal0% (1)
- Certificados Bol 491 PDFDocument38 pagesCertificados Bol 491 PDFMiguel José MárquezNo ratings yet
- Acontesim Ientos Upg: Visita de Plantel Cecyte VillagránDocument2 pagesAcontesim Ientos Upg: Visita de Plantel Cecyte VillagránIsmael FriasNo ratings yet
- Guía 6. Ciclo Celular - MitosisDocument12 pagesGuía 6. Ciclo Celular - MitosisBrian David Viana0% (1)
- 115 ADN ARN e Informacion PDFDocument14 pages115 ADN ARN e Informacion PDFTEDDY ARRIAGA BARRIOSNo ratings yet