You might also like
- Blood Vessels CH 13Document86 pagesBlood Vessels CH 13Nalla Mirelle CarbonellNo ratings yet
- Post Operative Nursing CareDocument5 pagesPost Operative Nursing CareJenjen Cortey100% (4)
- Daftar Isi Bridwell SpineDocument38 pagesDaftar Isi Bridwell Spineronny0% (1)
- Current and Future Development in Lung Cancer Diagnosis: Molecular SciencesDocument18 pagesCurrent and Future Development in Lung Cancer Diagnosis: Molecular SciencesHeru SigitNo ratings yet
- Simple Office SpirometryDocument42 pagesSimple Office SpirometryRoberto Merza III100% (4)
- Physical Diagnosis Case SampleDocument3 pagesPhysical Diagnosis Case SampleHerlene Lyneth Zalameda100% (2)
- Fontan Operation: DR Hussain BuxDocument62 pagesFontan Operation: DR Hussain BuxMujeeb Ur RehmanNo ratings yet
- Thoracic Decortication: Sabita Chalise Bns 3 YearDocument32 pagesThoracic Decortication: Sabita Chalise Bns 3 YearRamchandra Chalise100% (1)
- Retrieval Tools SystemsDocument3 pagesRetrieval Tools SystemsNithyaa SathishNo ratings yet
- Control System EbookDocument666 pagesControl System EbookNithyaa Sathish0% (1)
- CCD Guidelines 2023Document123 pagesCCD Guidelines 2023Cuong Nguyen Vu TheNo ratings yet
- Exploring The Berlin Definition of ArdsDocument40 pagesExploring The Berlin Definition of ArdsMuhammad RezaNo ratings yet
- Annual Update in Intensive Care and Emergency Medicine 2016Document488 pagesAnnual Update in Intensive Care and Emergency Medicine 2016Salvatore Cimino67% (6)
- 1996 Ite ExamDocument72 pages1996 Ite Exampiano543No ratings yet
- Preoperative Assessment and Premedication - PACUDocument60 pagesPreoperative Assessment and Premedication - PACUTraceNo ratings yet
- Correct answers for Filipino VI ReviewerDocument22 pagesCorrect answers for Filipino VI ReviewerYamSiriOdarnohNo ratings yet
- Congenital Heart Diseases - The Broken Heart - Clinical Features, Human Genetics and Molecular Pathways PDFDocument745 pagesCongenital Heart Diseases - The Broken Heart - Clinical Features, Human Genetics and Molecular Pathways PDFCamille Lyra de Espanha100% (1)
- Unrecognized Killers in EM Cardio NOTESDocument12 pagesUnrecognized Killers in EM Cardio NOTESawildstein100% (1)
- Essential anesthesia room setup checklistDocument31 pagesEssential anesthesia room setup checklistnbrassarNo ratings yet
- Regional 53Document53 pagesRegional 53RaMy “MhMd” ElaRabyNo ratings yet
- Control Systems Lecture Notes Laplace TransformDocument193 pagesControl Systems Lecture Notes Laplace Transformpduff_0675% (4)
- Echocardiographic Evaluation of Acute Aortic SyndromesDocument30 pagesEchocardiographic Evaluation of Acute Aortic SyndromesserubimNo ratings yet
- 2017 BMA Medical Book AwardsDocument45 pages2017 BMA Medical Book AwardsFerdinand Sta Ana Jr.No ratings yet
- Cardiac Defibrillation GÇô Mechanisms, Challenges and Implications PDFDocument260 pagesCardiac Defibrillation GÇô Mechanisms, Challenges and Implications PDFnaresh sharmaNo ratings yet
- Unit 1 - Cardiac Care UnitsDocument29 pagesUnit 1 - Cardiac Care UnitssitalekshmiNo ratings yet
- Sonography - 2014 - Cartwright - A Sonographer S Guide To The Assessment of Heart DiseaseDocument1 pageSonography - 2014 - Cartwright - A Sonographer S Guide To The Assessment of Heart Diseaseوجدان الدريبيNo ratings yet
- Primary Management in General, Vascular and Thoracic SurgeryDocument240 pagesPrimary Management in General, Vascular and Thoracic SurgeryCristhian Guevara CotrinaNo ratings yet
- JonasDocument561 pagesJonasАнастасияNo ratings yet
- General Principles of CirculationDocument52 pagesGeneral Principles of CirculationPhysiology by Dr RaghuveerNo ratings yet
- Pediatric Anesthesiology Review: Clinical Cases for Self-AssessmentFrom EverandPediatric Anesthesiology Review: Clinical Cases for Self-AssessmentNo ratings yet
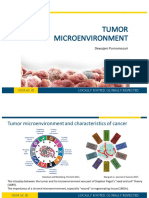
- 5 Tumor Micro EnvironmentDocument33 pages5 Tumor Micro Environmentanon_945728920No ratings yet
- Cardiac Anesthesia - Made Ridiculously Simple by Art Wallace, M.D., PH.DDocument17 pagesCardiac Anesthesia - Made Ridiculously Simple by Art Wallace, M.D., PH.Dstanwalks100% (1)
- Electrical TimeDocument14 pagesElectrical TimeNithyaa SathishNo ratings yet
- Training in Anaesthesia - Catherine SpoorsDocument4 pagesTraining in Anaesthesia - Catherine SpoorsgotufineNo ratings yet
- Anesthesia and Perioperative Care For Organ Transplantation-Kathirvel Subramaniam, Tetsuro Sakai (Eds.) - ASpringer-Verlag New York (2017) - Repaired PDFDocument572 pagesAnesthesia and Perioperative Care For Organ Transplantation-Kathirvel Subramaniam, Tetsuro Sakai (Eds.) - ASpringer-Verlag New York (2017) - Repaired PDFmarcoNo ratings yet
- Lecture 14-Oct.23 Haemodynamics of Circulation, Circulatory Mechanics, ResistanceDocument29 pagesLecture 14-Oct.23 Haemodynamics of Circulation, Circulatory Mechanics, ResistanceAparmitNo ratings yet
- (Studies in Mechanobiology, Tissue Engineering and Biomaterials 15) Thomas Franz (Eds.) - Cardiovascular and Cardiac Therapeutic Devices-Springer-Verlag Berlin Heidelberg (2014)Document246 pages(Studies in Mechanobiology, Tissue Engineering and Biomaterials 15) Thomas Franz (Eds.) - Cardiovascular and Cardiac Therapeutic Devices-Springer-Verlag Berlin Heidelberg (2014)Sheila Mae GardonNo ratings yet
- Dysrhythmia Interpretation Modules 1-6 June 2012Document126 pagesDysrhythmia Interpretation Modules 1-6 June 2012Jess Varose100% (3)
- Echocardiographic Assessment of Valve Stenosis: Heart Circulation Ultrasound SpecialistsDocument1 pageEchocardiographic Assessment of Valve Stenosis: Heart Circulation Ultrasound SpecialistsWayan GunawanNo ratings yet
- Capitulo 6Document19 pagesCapitulo 6gricelNo ratings yet
- Biofluid Mechanics - Exercises IIDocument2 pagesBiofluid Mechanics - Exercises IIMOnzerratheManzzanitaSixxNo ratings yet
- Blood Pressure Regulation SummaryDocument42 pagesBlood Pressure Regulation SummaryLouis JinNo ratings yet
- Cardio Lesson PlanDocument4 pagesCardio Lesson PlanAmiel Francisco Reyes100% (1)
- Tongue Diagnosis Maciocia OnlineDocument208 pagesTongue Diagnosis Maciocia OnlineCésar Ponce100% (1)
- Exam Questions HAAD-RN: HAAD Licensure Examination For Registered NursesDocument5 pagesExam Questions HAAD-RN: HAAD Licensure Examination For Registered NursesGreeshma victorNo ratings yet
- Copy-Question PapersDocument87 pagesCopy-Question Paperspooja patilNo ratings yet
- Lung Metabolism: Proteolysis and Antioproteolysis Biochemical Pharmacology Handling of Bioactive SubstancesFrom EverandLung Metabolism: Proteolysis and Antioproteolysis Biochemical Pharmacology Handling of Bioactive SubstancesAlain JunodNo ratings yet
- Cardiology Ultrasound EmergencyDocument92 pagesCardiology Ultrasound Emergencybeeluvit81444No ratings yet
- AHA Statement-Cardiopulmonary Impact of E-Cigarettes and Vaping-2023Document26 pagesAHA Statement-Cardiopulmonary Impact of E-Cigarettes and Vaping-2023Walter ReyesNo ratings yet
- Oliguria 6oct2010 FinalDocument75 pagesOliguria 6oct2010 Finalsallam8008No ratings yet
- Immediate Complications Following Thoracic SurgeryDocument17 pagesImmediate Complications Following Thoracic SurgeryPopa TiberiuNo ratings yet
- Acute Myocardial Ischaemia ESICMDocument76 pagesAcute Myocardial Ischaemia ESICMpaskariatne probo dewiNo ratings yet
- The Perfusion Crisis ManualDocument3 pagesThe Perfusion Crisis Manualghg sddNo ratings yet
- Recruitment Maneuvers and PEEP TitrationDocument17 pagesRecruitment Maneuvers and PEEP TitrationDiego Cruces OrdoñezNo ratings yet
- NEJM 2014 Fundamentals of Lung AuscultationDocument7 pagesNEJM 2014 Fundamentals of Lung AuscultationFelipe CeaNo ratings yet
- AGT Applied Anatomy Notes1.1Document69 pagesAGT Applied Anatomy Notes1.1Akshay ChauhanNo ratings yet
- Miller's Anesthesia, 8th Edition: Anesthesiology April 2016Document3 pagesMiller's Anesthesia, 8th Edition: Anesthesiology April 2016Sabbir AhammedNo ratings yet
- Should We Use Driving Pressure To Set Tidal Volume? PDFDocument7 pagesShould We Use Driving Pressure To Set Tidal Volume? PDFYaxkin NikNo ratings yet
- Pop Cardiac Surgery Critical Care 2015Document20 pagesPop Cardiac Surgery Critical Care 2015Eward Rod SalNo ratings yet
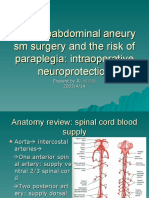
- Thoracoabdominal Aneurysm Surgery and The Risk of ParaplegiaDocument34 pagesThoracoabdominal Aneurysm Surgery and The Risk of ParaplegiaKhaled S. HarbNo ratings yet
- Harvesting The Radial ArteryDocument11 pagesHarvesting The Radial ArteryprofarmahNo ratings yet
- STRUCTURE AND FUNCTION OF THE HEARTDocument16 pagesSTRUCTURE AND FUNCTION OF THE HEARTPkernNo ratings yet
- Cardiac AnatomyDocument6 pagesCardiac Anatomysandranamahen2No ratings yet
- 9780190685157Document639 pages9780190685157Jaber AlhaizaeyNo ratings yet
- CCTGA Diagnosis, Treatment & OutcomesDocument94 pagesCCTGA Diagnosis, Treatment & OutcomesVik SharNo ratings yet
- Anatomical Guide For ElectromyographerDocument397 pagesAnatomical Guide For ElectromyographerBelén GonzálezNo ratings yet
- CARDIAC ELECTROPHYSIOLOGY AND THE ELECTROCARDIOGRAM - ClinicalKey PDFDocument40 pagesCARDIAC ELECTROPHYSIOLOGY AND THE ELECTROCARDIOGRAM - ClinicalKey PDFMelanie DascăluNo ratings yet
- 1) Vasoplegia During Cardiac Surgery Current Concepts and Management 2010Document5 pages1) Vasoplegia During Cardiac Surgery Current Concepts and Management 2010Carolina QuirogaNo ratings yet
- Non Cardiac OpDocument21 pagesNon Cardiac Opc4ri5No ratings yet
- Acid Base PhysiologyDocument1 pageAcid Base PhysiologyPrabjot SehmiNo ratings yet
- CPBDocument48 pagesCPBRaghavendra PrasadNo ratings yet
- Failed Spinal Anesthesia PDFDocument2 pagesFailed Spinal Anesthesia PDFShamim100% (1)
- David Gozal's CVDocument87 pagesDavid Gozal's CVMSKCNo ratings yet
- Lecture 9 BurnsDocument4 pagesLecture 9 BurnsRose Ann RaquizaNo ratings yet
- 1 s2.0 S1607551X11001835 MainDocument13 pages1 s2.0 S1607551X11001835 MainNithyaa SathishNo ratings yet
- Design of Artificial Arms and Hands For Prosthetic ApplicationsDocument28 pagesDesign of Artificial Arms and Hands For Prosthetic ApplicationsNithyaa SathishNo ratings yet
- Types of WheelchairsDocument13 pagesTypes of WheelchairsMehnaz KhanNo ratings yet
- EGGDocument14 pagesEGGNithyaa SathishNo ratings yet
- Hilbert-Huang Transform and Its Applications in Engineering and Biomedical Signal AnalysisDocument8 pagesHilbert-Huang Transform and Its Applications in Engineering and Biomedical Signal AnalysisNithyaa SathishNo ratings yet
- New Electric Prosthetic System with Simultaneous Elbow & Hand ControlDocument10 pagesNew Electric Prosthetic System with Simultaneous Elbow & Hand ControlNithyaa SathishNo ratings yet
- Diabetic Foot Ulcers - Prevention, Diagnosis and Classification - American Family PhysicianDocument6 pagesDiabetic Foot Ulcers - Prevention, Diagnosis and Classification - American Family PhysicianNithyaa SathishNo ratings yet
- Telehealth (MX9014)Document2 pagesTelehealth (MX9014)Nithyaa SathishNo ratings yet
- Encryption MatlabDocument9 pagesEncryption MatlabKaris Shang100% (1)
- Part BDocument2 pagesPart BNithyaa SathishNo ratings yet
- Pattern NotesDocument23 pagesPattern NotesArchanaBmeNo ratings yet
- ElectronicsDocument8 pagesElectronicsNithyaa SathishNo ratings yet
- Webb CHPT 6s4Document14 pagesWebb CHPT 6s4aloove66No ratings yet
- Radiopharmaceuticals For Radioisotope ImagingDocument21 pagesRadiopharmaceuticals For Radioisotope ImagingNithyaa SathishNo ratings yet
- Automated Detection and Classification of Parasitic and Non-Parasitic DiseasesDocument12 pagesAutomated Detection and Classification of Parasitic and Non-Parasitic DiseasesNithyaa SathishNo ratings yet
- ch54MEASUREMENT TECHNIQUES IN RESPIRATORY MECHANICSDocument16 pagesch54MEASUREMENT TECHNIQUES IN RESPIRATORY MECHANICSNithyaa SathishNo ratings yet
- Mathematical Modeling of Respiratory System: A ReviewDocument5 pagesMathematical Modeling of Respiratory System: A ReviewNithyaa SathishNo ratings yet
- Bonus Chapt 07Document36 pagesBonus Chapt 07Niki KarmannNo ratings yet
- As 04 Transport in Animals Ans - BiologyDocument7 pagesAs 04 Transport in Animals Ans - BiologyUncleBulgaria100% (1)
- General Heart Diagram, Handout Heart Diagram, Vessels HandoutDocument4 pagesGeneral Heart Diagram, Handout Heart Diagram, Vessels HandoutJNo ratings yet
- Transportation in Animals and Plants: BloodDocument12 pagesTransportation in Animals and Plants: BloodSeiko classesNo ratings yet
- Body Fluids and CirculationsDocument14 pagesBody Fluids and CirculationsNikhil SharmaNo ratings yet
- Life Processes Class 10 Notes - Transportation in Humans & PlantsDocument4 pagesLife Processes Class 10 Notes - Transportation in Humans & PlantsGeetanjali jadhavNo ratings yet
- Grade 7Document6 pagesGrade 7gnanabalanNo ratings yet
- How to perform the Palm Tree Pose (TalasanaDocument8 pagesHow to perform the Palm Tree Pose (Talasanaabhilasha_yadav_1No ratings yet
- Chapter 11Document17 pagesChapter 112071317No ratings yet
- Physiology of PregnancyDocument5 pagesPhysiology of PregnancyAlejandra RequesensNo ratings yet
- Study of Retinal Vessel Oxygen Saturation in Ischemic and Non-Ischemic Branch Retinal Vein OcclusionDocument9 pagesStudy of Retinal Vessel Oxygen Saturation in Ischemic and Non-Ischemic Branch Retinal Vein OcclusionEvenjelina EveNo ratings yet
- Emailing Diabetic Wound Care White PaperDocument20 pagesEmailing Diabetic Wound Care White PaperGreatAkbar1No ratings yet
- Chapter 11 The Cardiovascular SystemDocument25 pagesChapter 11 The Cardiovascular SystemYeshia InocencioNo ratings yet
- IGCSE - Bio - Lesson Plan 5 - Blood and CirculationDocument3 pagesIGCSE - Bio - Lesson Plan 5 - Blood and CirculationHisokagenNo ratings yet
- Elektro Fisika Dan Sumber Fisis 1 Pertemuan 2Document21 pagesElektro Fisika Dan Sumber Fisis 1 Pertemuan 2estherNo ratings yet
- Overview of FistulasDocument39 pagesOverview of FistulasWade JacksonNo ratings yet
- How to Treat Varicose Veins (39Document2 pagesHow to Treat Varicose Veins (39Tee VillanuevaNo ratings yet
- Circulatory System REVIEWERDocument6 pagesCirculatory System REVIEWERClyde BaltazarNo ratings yet
- Chapter 7 BehaviorDocument14 pagesChapter 7 BehaviorVanny RamosNo ratings yet
- Fibula Free Flap 2Document5 pagesFibula Free Flap 2Sanjay KumarNo ratings yet
- A Short Bevel Needle With A Very Thin TipDocument8 pagesA Short Bevel Needle With A Very Thin Tipmohammadreza hajialiNo ratings yet