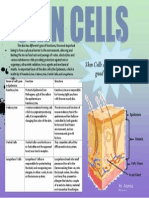
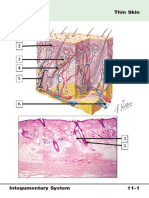
You might also like
- From Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.From EverandFrom Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.No ratings yet
- Pdia 30 20287Document12 pagesPdia 30 20287angelos96papNo ratings yet
- Transferencia de MelaninaDocument11 pagesTransferencia de MelaninaEridani OlmosNo ratings yet
- IjmsDocument20 pagesIjmsNikhola RisolNo ratings yet
- Chapter 72:: Biology of Melanocytes:: Hee-Young Park & Mina YaarDocument16 pagesChapter 72:: Biology of Melanocytes:: Hee-Young Park & Mina YaarMeutiDonytaNo ratings yet
- The Regulation of Skin Pigmentation: MinireviewDocument5 pagesThe Regulation of Skin Pigmentation: MinireviewMar Kristian Vero LumenNo ratings yet
- The Melanocyte Lineage in Development and DiseaseDocument13 pagesThe Melanocyte Lineage in Development and DiseaseLuis Kamil Buitrago OsornoNo ratings yet
- Yamaguchi 2009Document7 pagesYamaguchi 2009Gabriela LpezNo ratings yet
- Artikel 2 Regulation of Skin PigmentationDocument5 pagesArtikel 2 Regulation of Skin PigmentationNandhy AlpNo ratings yet
- Torello Lotti, MD: Professor of Dermatology and Venereology Florence, ItalyDocument54 pagesTorello Lotti, MD: Professor of Dermatology and Venereology Florence, ItalyBrîndușa PetruțescuNo ratings yet
- Uyen2008 Article MechanismOfSkinPigmentationDocument14 pagesUyen2008 Article MechanismOfSkinPigmentationBungaNo ratings yet
- The Biology of Melanocytes (Pages 57-65)Document9 pagesThe Biology of Melanocytes (Pages 57-65)jenNo ratings yet
- Dermatology Questions and AnsDocument147 pagesDermatology Questions and AnsEl FaroukNo ratings yet
- Biomolecules 12 01874 PDFDocument20 pagesBiomolecules 12 01874 PDFAhmad TaufikNo ratings yet
- Structure and Function of Melanocytes: Microscopic Morphology and Cell Biology of Mouse Melanocytes in The Epidermis and Hair FollicleDocument15 pagesStructure and Function of Melanocytes: Microscopic Morphology and Cell Biology of Mouse Melanocytes in The Epidermis and Hair FollicleRog DonNo ratings yet
- Structure & Funtion NailsDocument5 pagesStructure & Funtion NailsrikaNo ratings yet
- Pertumbuhan SelDocument37 pagesPertumbuhan SelbeniNo ratings yet
- Anatomy, Skin (Integument), Epidermis - StatPearls - NCBI BookshelfDocument8 pagesAnatomy, Skin (Integument), Epidermis - StatPearls - NCBI Bookshelfpka25No ratings yet
- Signaling Pathways in Melanogenesis: Molecular SciencesDocument18 pagesSignaling Pathways in Melanogenesis: Molecular SciencesEllen FernandaNo ratings yet
- Ijms-21-09769-V2Document18 pagesIjms-21-09769-V2Nikhola RisolNo ratings yet
- UV Radiation and Skin PDFDocument27 pagesUV Radiation and Skin PDFDesi RatnaningtyasNo ratings yet
- Cs 1Document1 pageCs 1api-223339668No ratings yet
- Curso de La PielDocument11 pagesCurso de La PielNicole ReyesNo ratings yet
- 1,2-Introduction To DermatologyDocument20 pages1,2-Introduction To Dermatologyhesham attiaNo ratings yet
- Lecture 27 - Histology of The Integumentary System 2Document89 pagesLecture 27 - Histology of The Integumentary System 2spitzmark2030No ratings yet
- M13 v2 Dermatology & STI - LO3Document8 pagesM13 v2 Dermatology & STI - LO3medipro clinicNo ratings yet
- NIACINAMIDE 5% BrighteningDocument12 pagesNIACINAMIDE 5% Brighteningproduksi roiNo ratings yet
- Skin Structure and Function: Applied Dermatotoxicology. DOI: © 2014 Elsevier Inc. All Rights ReservedDocument10 pagesSkin Structure and Function: Applied Dermatotoxicology. DOI: © 2014 Elsevier Inc. All Rights ReservedArsene AngelaNo ratings yet
- Histophysiology+of+the+Skin+for+Vula+ Dr+MowlaDocument28 pagesHistophysiology+of+the+Skin+for+Vula+ Dr+MowlaYaaseen GallantNo ratings yet
- Cosmetics 08 00082 v2Document11 pagesCosmetics 08 00082 v2laithuyduong007No ratings yet
- Safari - Feb 21, 2024 at 11:48 AMDocument1 pageSafari - Feb 21, 2024 at 11:48 AMsyansyncNo ratings yet
- Integumentary SystemDocument24 pagesIntegumentary SystemWadabiNo ratings yet
- Meth Elab Topic N1Document23 pagesMeth Elab Topic N1Group M1869No ratings yet
- Skin Basic Structure and FunctionDocument11 pagesSkin Basic Structure and FunctionPanggih Sekar Palupi IINo ratings yet
- Transcription Physiology'' of Pigment Formation in Melanocytes: Central Role of MITFDocument11 pagesTranscription Physiology'' of Pigment Formation in Melanocytes: Central Role of MITFAbdur Rachman Ba'abdullahNo ratings yet
- Biology of Melanocytes: Done By: Mohammed Abduljabbar Done By: Mohammed Abduljabbar KauhDocument66 pagesBiology of Melanocytes: Done By: Mohammed Abduljabbar Done By: Mohammed Abduljabbar Kauhmoon1312No ratings yet
- Skin AnatomyDocument71 pagesSkin AnatomyHardik DaveNo ratings yet
- SkinDocument20 pagesSkinDr. Mary MjeedNo ratings yet
- Complex Networks Orchestrate Epithelial-Mesenchymal Transitions (Σημειωμένο)Document12 pagesComplex Networks Orchestrate Epithelial-Mesenchymal Transitions (Σημειωμένο)panna1No ratings yet
- MelanocytesDocument3 pagesMelanocytesأحمد المسيريNo ratings yet
- Ujdvc 0121Document9 pagesUjdvc 0121Christian DiehlNo ratings yet
- Anatomy Histo Physio DR - AdilDocument73 pagesAnatomy Histo Physio DR - AdilHybat ElsheikhNo ratings yet
- Integumentary System: Stratum BasaleDocument21 pagesIntegumentary System: Stratum BasalerinaNo ratings yet
- Histology SkinDocument5 pagesHistology SkinFahd Abdullah Al-refaiNo ratings yet
- "Vision Without Action Is A Daydream. Action Without Vision Is A Nightmare." Japanese ProverbDocument22 pages"Vision Without Action Is A Daydream. Action Without Vision Is A Nightmare." Japanese Proverb123456100% (1)
- Fits Patrick Chap 20Document8 pagesFits Patrick Chap 20FIT ChannelNo ratings yet
- Bab 7. Struktur KulitDocument56 pagesBab 7. Struktur KulityumizoneNo ratings yet
- Lecture Presented by:-ALI Waheed /yasmin Falah /zainab Mazin Supervised By:-Dr. Sattar Jabbar Jasim /cell and Tissue BiomedicalDocument24 pagesLecture Presented by:-ALI Waheed /yasmin Falah /zainab Mazin Supervised By:-Dr. Sattar Jabbar Jasim /cell and Tissue BiomedicalAli Waheed jolan100% (1)
- The Following Notes Are References According To Figures and Tables in Gartner and Hiatt (2 Edition, 2001)Document5 pagesThe Following Notes Are References According To Figures and Tables in Gartner and Hiatt (2 Edition, 2001)gerginNo ratings yet
- Schwartz's Principles of Surgery: CH16: Page 1Document13 pagesSchwartz's Principles of Surgery: CH16: Page 1Kathryn ReunillaNo ratings yet
- UC Davis: Dermatology Online JournalDocument7 pagesUC Davis: Dermatology Online JournalFerdie YuzonNo ratings yet
- Lit SkinStruct Bensouillah Ch01 PDFDocument11 pagesLit SkinStruct Bensouillah Ch01 PDFisaco1531012No ratings yet
- Relation Between Hypothermia and SSIDocument7 pagesRelation Between Hypothermia and SSIRekkiNo ratings yet
- What Are The Properties of Normal SkinDocument102 pagesWhat Are The Properties of Normal Skinlovely_dyaNo ratings yet
- How, and From Which Cell Sources, Do Nevi Really Develop?Document19 pagesHow, and From Which Cell Sources, Do Nevi Really Develop?Brîndușa PetruțescuNo ratings yet
- 1.integumentary System - HistologyDocument34 pages1.integumentary System - Histologyqty9jgkpnzNo ratings yet
- Transplantation of Melanocyte Stem Cells in Vitiliginous SkinDocument12 pagesTransplantation of Melanocyte Stem Cells in Vitiliginous SkinririnNo ratings yet
- Keratinocytes Undergo A Differentiation Process, Starting From A Single Layer of Stem Cells at The Basis ofDocument14 pagesKeratinocytes Undergo A Differentiation Process, Starting From A Single Layer of Stem Cells at The Basis ofHarish KakraniNo ratings yet
- Bouncing Checks: Liabilities For Violation of B.P. 22Document10 pagesBouncing Checks: Liabilities For Violation of B.P. 22Nickford AcidoNo ratings yet
- Estate TaxationDocument9 pagesEstate TaxationJanelleNo ratings yet
- Law Prelim ReviewerDocument5 pagesLaw Prelim ReviewerJanelleNo ratings yet
- Advacc 2 Conso. Fs-Subsequent To Date of AcquisitionDocument11 pagesAdvacc 2 Conso. Fs-Subsequent To Date of AcquisitionJanelleNo ratings yet
- Bouncing Checks: Liabilities For Violation of B.P. 22Document10 pagesBouncing Checks: Liabilities For Violation of B.P. 22Nickford AcidoNo ratings yet
- GOVERNMENT ACCOUNTING - Accounting Responsibilities.Document19 pagesGOVERNMENT ACCOUNTING - Accounting Responsibilities.Hannah Verano86% (7)
- 217 Facts About BTS Ebook PDFDocument16 pages217 Facts About BTS Ebook PDFJanelleNo ratings yet
- Property DisposalDocument2 pagesProperty DisposalJanelleNo ratings yet
- Who Bears The Cost of Excessive Executive Compensation (And OtherDocument23 pagesWho Bears The Cost of Excessive Executive Compensation (And OtherJanelleNo ratings yet
- ch01 2accDocument11 pagesch01 2accronin214No ratings yet
- Southwest Airlines A Rough Patch or Permanent DescentDocument10 pagesSouthwest Airlines A Rough Patch or Permanent DescentJanelleNo ratings yet
- The Collapse of EnronDocument9 pagesThe Collapse of EnronJanelleNo ratings yet
- Chapter 1-Introduction To Managerial Accounting: True/FalseDocument15 pagesChapter 1-Introduction To Managerial Accounting: True/FalseJanelleNo ratings yet
- 217 Facts About BTS Ebook PDFDocument16 pages217 Facts About BTS Ebook PDFJanelleNo ratings yet
- Southwest Airlines 2002 An Industry Under SiegeDocument8 pagesSouthwest Airlines 2002 An Industry Under SiegeJanelle50% (2)
- Social Science Theories NotesDocument1 pageSocial Science Theories NotesJanelleNo ratings yet
- Ais Chapter 15 Rea ModelDocument138 pagesAis Chapter 15 Rea ModelJanelleNo ratings yet
- Social Science Theories NotesDocument1 pageSocial Science Theories NotesJanelleNo ratings yet
- Business Policy and Strategy NotesDocument1 pageBusiness Policy and Strategy NotesJanelleNo ratings yet
- Compilation of Problems in FactoringDocument9 pagesCompilation of Problems in FactoringJanelleNo ratings yet
- Respiratory & Circulatory SystemDocument6 pagesRespiratory & Circulatory SystemJanelleNo ratings yet
- Information Technology in Internal AuditingDocument1 pageInformation Technology in Internal AuditingJanelleNo ratings yet
- Ratinality of Growing Sugar in NegrosDocument31 pagesRatinality of Growing Sugar in NegrosJanelleNo ratings yet
- Work and Power Practice 2 AnswersDocument4 pagesWork and Power Practice 2 AnswersJanelleNo ratings yet
- Eliminating Sentence FragmentsDocument1 pageEliminating Sentence FragmentsJanelleNo ratings yet
- Keratinocytes KeratinDocument16 pagesKeratinocytes KeratinJanelleNo ratings yet
- Cse Integration Lesson PlanDocument5 pagesCse Integration Lesson PlanCarl Anthony Lague PahuyoNo ratings yet
- Frog WorksheetDocument4 pagesFrog WorksheetCrystal Joy RebelloNo ratings yet
- PNU Final Coaching Gen - EdDocument15 pagesPNU Final Coaching Gen - EdTatinNo ratings yet
- Designing For The Human SensesDocument50 pagesDesigning For The Human SensesRahaf MachaalNo ratings yet
- The Lab Session Report of Biological Techniques in Laboratory and Field (BIO403) Animals and Plants Colllection TechniquesDocument5 pagesThe Lab Session Report of Biological Techniques in Laboratory and Field (BIO403) Animals and Plants Colllection TechniquesUmmi SalamahNo ratings yet
- Effect of Square Stepping Exercise Versus Swiss Ball Exercise On Balance in Institutionalized Elderly PopulationDocument6 pagesEffect of Square Stepping Exercise Versus Swiss Ball Exercise On Balance in Institutionalized Elderly Populationastrinila fauziNo ratings yet
- Let Drills Gen Ed General ScienceDocument132 pagesLet Drills Gen Ed General ScienceJoyanne DiwaNo ratings yet
- BiologyDocument41 pagesBiologymina0% (1)
- Astriana Final MRM Project Paper-RevisedDocument92 pagesAstriana Final MRM Project Paper-RevisedNgurah MahasviraNo ratings yet
- Energy Pyramid - Activity StephanieDocument4 pagesEnergy Pyramid - Activity Stephanieapi-473892313No ratings yet
- Wed09 NovozymesDocument29 pagesWed09 NovozymesPulbere NeagraNo ratings yet
- UPCAT Complete Reviewer 2020jjjjjjDocument84 pagesUPCAT Complete Reviewer 2020jjjjjjPolyblank Glitch100% (2)
- Applied Zoology: Pearl Culture Bechan LalDocument16 pagesApplied Zoology: Pearl Culture Bechan LalSk RajNo ratings yet
- Impact Factor Journals 2008Document166 pagesImpact Factor Journals 2008ghassemikermanNo ratings yet
- Answers For Support Worksheet - Option F: 1 A Feature Bacteria Archaea EukaryotaDocument1 pageAnswers For Support Worksheet - Option F: 1 A Feature Bacteria Archaea EukaryotaHellö PëachNo ratings yet
- Sunrise Sunrise: zb1) Z b2)Document26 pagesSunrise Sunrise: zb1) Z b2)jn875849No ratings yet
- BAM Chapter 29. CronobacterDocument14 pagesBAM Chapter 29. CronobacterremyNo ratings yet
- Midterm 3TDocument10 pagesMidterm 3TThanh JessieNo ratings yet
- Long Range Sports Med 15Document6 pagesLong Range Sports Med 15api-361293242No ratings yet
- Minimal Intervention Dentistry in General Dentistry PDFDocument10 pagesMinimal Intervention Dentistry in General Dentistry PDFmostafahassan777No ratings yet
- Strategies For Plant Regeneration From Mesophyll Protoplasts of The Recalcitrant Fruit and Farmwoodland Species L. (Sweet/wild Cherry)Document6 pagesStrategies For Plant Regeneration From Mesophyll Protoplasts of The Recalcitrant Fruit and Farmwoodland Species L. (Sweet/wild Cherry)Geral BlancoNo ratings yet
- The Case For Increased Research On Male FertilityDocument23 pagesThe Case For Increased Research On Male FertilityIndyra AlencarNo ratings yet
- Oil EOR HandbookDocument24 pagesOil EOR HandbookRicardo Hernan Gomez BNo ratings yet
- Arndt Schmidt-Denter Auer Weitere 2003Document15 pagesArndt Schmidt-Denter Auer Weitere 2003Louis Fetilo FabunanNo ratings yet
- Components of Physical FitnessDocument1 pageComponents of Physical Fitnessarjay zxcNo ratings yet
- Introduction To Genetic Algorithms (GA)Document14 pagesIntroduction To Genetic Algorithms (GA)Somya AryaNo ratings yet
- Dalandan (Citrus Aurantium Linn.) Peelings and Calamansi (Citrofortunella Microcarpa) Extract As An Alternative Fire Ant KillerDocument6 pagesDalandan (Citrus Aurantium Linn.) Peelings and Calamansi (Citrofortunella Microcarpa) Extract As An Alternative Fire Ant KillerMaryknoll BernabeNo ratings yet
- Photosynthesis and RespiratDocument12 pagesPhotosynthesis and Respiratarlyn09160% (2)
- BRO Fluorescence SpectrosDocument24 pagesBRO Fluorescence SpectrosWaleed El-azabNo ratings yet
- Photoexcited Molecules of Pteridine DerivativesDocument6 pagesPhotoexcited Molecules of Pteridine DerivativesDr. Amrit MitraNo ratings yet
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (4)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (517)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (393)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (411)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (812)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (109)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (595)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (216)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)